
Climate Change Impacts on the Marine Cycling of Biogenic Sulfur: A Review


1
Coasts and Ocean Research, Oceans and Atmosphere, CSIRO, Canberra, ACT 2601, Australia
2
School of Environment and Science, Griffith University, Nathan, QLD 4111, Australia
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(8), 1581; https://doi.org/10.3390/microorganisms10081581
Submission received: 29 June 2022
/
Revised: 2 August 2022
/
Accepted: 2 August 2022
/
Published: 5 August 2022
(This article belongs to the Special Issue Phytoplankton-Bacteria Interactions 2.0)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:A key component of the marine sulfur cycle is the climate-active gas dimethylsulfide (DMS), which is synthesized by a range of organisms from phytoplankton to corals, and accounts for up to 80% of global biogenic sulfur emissions. The DMS cycle starts with the intracellular synthesis of the non-gaseous precursor dimethylsulfoniopropionate (DMSP), which is released to the water column by various food web processes such as zooplankton grazing. This dissolved DMSP pool is rapidly turned over by microbially mediated conversion using two known pathways: demethylation (releasing methanethiol) and cleavage (producing DMS). Some of the formed DMS is ventilated to the atmosphere, where it undergoes rapid oxidation and contributes to the formation of sulfate aerosols, with the potential to affect cloud microphysics, and thus the regional climate. The marine phase cycling of DMS is complex, however, as heterotrophs also contribute to the consumption of the newly formed dissolved DMS. Interestingly, due to microbial consumption and other water column sinks such as photolysis, the amount of DMS that enters the atmosphere is currently thought to be a relatively minor fraction of the total amount cycled through the marine food web—less than 10%. These microbial processes are mediated by water column temperature, but the response of marine microbial assemblages to ocean warming is poorly characterized, although bacterial degradation appears to increase with an increase in temperature. This review will focus on the potential impact of climate change on the key microbially mediated processes in the marine cycling of DMS. It is likely that the impact will vary across different biogeographical regions from polar to tropical. For example, in the rapidly warming polar oceans, microbial communities associated with the DMS cycle will likely change dramatically during the 21st century with the decline in sea ice. At lower latitudes, where corals form an important source of DMS (P), shifts in the microbiome composition have been observed during thermal stress with the potential to alter the DMS cycle.
1. Introduction
Dimethylsulfide (DMS: CH3SCH3) is the most abundant form of volatile sulfur (S) in the ocean and is the main biogenic source of reduced S to the global atmosphere [1]. First Shaw (1983) [2] and then Charlson et al. [3] posited links between DMS emission to the atmosphere and the formation of atmospheric sulfate aerosols potentially affecting global climate. It was hypothesized that a temperature-driven change in marine phytoplankton growth would increase DMS emissions to the atmosphere, leading to an increase in sulfate aerosol, potentially forming more cloud condensation nuclei (CCN) and brighter clouds. This change in cloud microphysics could cool the earth’s surface and thus stabilize the climate against perturbations due to greenhouse gas warming. The proposed DMS–climate link, later called the “CLAW hypothesis” (the acronym is based on the names of the authors of the Charlson et al. [3] paper), stimulated a flurry of research in subsequent decades [4], with over 17,000 hits on the phrase “dimethylsulfide and climate” in a 2022 search on Google Scholar. The multi-layered complexity of CLAW generated research in a variety of previously unconnected and disparate disciplines (e.g., atmospheric chemistry and marine biology) and can be considered an early motivation for the development of the field of Earth System Science. Notwithstanding an enormous research effort over the last three decades, there is still no general consensus on the global validity of the original CLAW hypothesis [5,6].
The sea-to-air flux of sulfur due to DMS is estimated to be in the range of (18–31) Tg S year−1 [7], which constitutes about 50% of the total biogenic atmospheric sulfate burden [8,9]. In comparison, anthropogenic sulfur emissions dominate the global sulfur flux [10], and the formation of anthropogenic aerosol has caused a cooling of the global climate during the industrial period, partly offsetting the greenhouse gas warming [11]. However, this dominance of anthropogenic over natural aerosol may change during the 21st century (C) as more rigorous air pollution controls are implemented [12], with negative trends in Europe and North America already evident [13].
Once ventilated to the atmosphere, DMS is rapidly oxidized to form non-sea-salt sulfate (nss-SO42−) and methanesulfonate (MSA) aerosols, but the efficiency of this process in producing CCN has been questioned [14]. The atmospheric chemistry of DMS is complex and still not fully understood, with novel oxidation pathways and new byproducts still being discovered [15,16].
Notwithstanding the doubts about the CLAW hypothesis, ongoing research is uncovering new ecological roles for DMS. Various marine organisms can produce intracellular dimethylsulfoniopropionate (DMSP), the precursor compound to DMS. It has been long recognized that different phytoplankton genera synthesize DMSP to varying degrees [17], with coccolithophorids and small flagellates having higher intracellular concentrations of DMSP, which was originally thought to act as an osmolyte in the algal cell. Further research has identified several other roles for DMSP, such as a response to oxidative stress in algae and corals [18], and giant clams [19], and as an osmoregulant and reduced carbon/sulfur source in marine heterotrophic bacteria [20,21,22]. In addition, DMS can act as a scavenger of reactive oxygen species (ROS) via conversion to dimethylsulfoxide (DMSO) [18,23]. Although phytoplankton have long been recognized as synthesizers of DMSP, other marine organisms such as benthic and pelagic macro-algae, corals, and sponges have also been identified as important sources of DMSP [24,25,26,27].
Recent observational studies have confirmed the importance of marine DMS emissions to atmospheric CCN [28,29] and non-sea-salt aerosol burden [30]. Additionally, strong broadscale correlations between DMS concentration and solar radiation have also been reported [31] and between solar radiation and DMS synthesis [32]. Overall, these studies lend support to parts of the CLAW hypothesis, however modeling attempts to assess the direction and magnitude of the DMS–climate feedback [33,34,35] have often led to contradictory results [36,37,38,39,40]. Other regional modeling studies indicate significant meridional variability in future DMS emissions under warming, with the strongest response simulated at high latitudes in both hemispheres [41,42,43,44].
Marine bacteria were first recognized for their role in the decomposition of organic material and the remineralization of inorganic nutrients, a role that only became fully understood during the 1980s. Azam and colleagues described a “microbial loop” in which dissolved organic matter released by phytoplankton and grazers is consumed by bacteria. Bacteria are consumed by protozoa, which are then consumed by microzooplankton that are part of the traditional grazing chain [45]. It was soon also understood that microbial organisms played key roles in the biogeochemistry of the ocean [46]. This is evidenced by the fact that the entire pelagic microbial food web, including protozoan microzooplankton, is typically 5–10 times the mass of all multicellular aquatic organisms, namely, zooplankton and fish [47].
However, the role of microbial heterotrophs in the DMS (P) cycle was not fully appreciated at the time of the original CLAW hypothesis. Indeed, it was not until two seminal papers by Simo [48] and Kiene et al. [49] that the complex role of microbes in the cycling of DMS (P) began to be better understood. The marine sulfur cycle is now considered a quintessential example of algal–bacterial interaction [50], and is the focus of intense research interest, with new components and pathways still being identified [51].
Algal intracellular DMSP can be released to the water column via a number of processes, namely, zooplankton grazing [52], exudation [53], viral lysis [54], or cell senescence. It is now well-understood that bacterioplankton are key agents in the consumption of dissolved DMSP [20], the conversion of DMSP to DMS [55], and also the consumption of DMS [56,57]. Less well-understood is the impact of elevated CO2 and warming on the marine microbial loop and the cycling of DMS (P). As noted by Cavicchioli et al. [58], the reciprocal effects of microorganisms on climate change and climate change on microorganisms is complex and our understanding is still incomplete. This review will examine the potential for climate change to impact on the marine bacterioplankton community structure and function, and what this may mean for future DMS (P) synthesis and emissions.
2. The Marine DMS Cycle
It is now recognized that organosulfur compounds both in the water and the cell play important roles in the marine food web by mediating interactions between phytoplankton and heterotrophic bacteria [59,60]. Sulfur is an essential element for marine primary production, with its algal cellular stoichiometry similar to phosphorus [61]. Thus, marine phytoplankton play a key role in the global S cycle because they can assimilate sulfate (SO42−), which is abundant in the ocean. Sea-to-air transfer of S occurs via the ability of phytoplankton to synthesize cellular DMSP, the microbial-mediated transformation to DMS, and its subsequent emission to the atmosphere, where it has a possible involvement in climate regulation. As noted above, DMS provides the main biogenic source of reduced S to the global atmosphere. The annual global production of DMSP by deep ocean phytoplankton (excluding coastal) is conservatively estimated as 3.8 Pg C year−1 [62]—this is a substantial amount for a single compound, equal to approximately 10% of the total carbon fixed annually by marine phytoplankton [63]. DMSP is also critical to the structure of the marine food web as it provides a substantial source of reduced carbon and sulfur for heterotrophic bacteria [49,64] and can also be concentrated internally to serve as an osmolyte, reaching concentrations of 70 mM in some marine bacteria [21,65]. Field studies indicate that particulate DMSP concentration can span a very large range depending on the phytoplankton speciation and bloom dynamics, from 5 to >4000 nM [66,67], whereas dissolved DMSP is often present in lower concentrations (1–25 nM) and has a turnover rate of 1–129 nM d−1 [49].
2.1. Key Food Web Processes
A conceptual model of the key processes in the marine cycle of DMS and its precursor compound DMSP is presented in Figure 1. Both these compounds can occur in particulate and dissolved phases. The processes included are those described for an algal DMSP source by Stefels et al. [68] and Kiene et al. [49], and emphasize the central role of heterotrophic bacteria. The main bacteria-mediated processes are colored red (Figure 1). They include the conversion of dissolved DMSP to methanethiol (MeSH; CH3SH), via the demethiolation pathway, or to DMS via the enzymatic cleavage pathway, and the consumption of both dissolved DMSP and DMS by bacteria. Interestingly, MeSH is also a volatile compound but its sea-to-air flux estimates are scant, although it has recently been estimated to be comparable to, albeit less than, that of DMS [69].
However, this conceptual model is only partly complete as Li et al. [70] describe a non-bioavailable pool (refractory) of dissolved DMSP, which they accounted for by retention in bacteria, but also partly unknown. Intriguingly, Liu et al. [71] report the synthesis of DMSP by bacteria in deep aphotic waters or surficial sediments, suggesting that this process is largely independent of phytoplankton production, although detrital DMSP from zooplankton grazing could also be a source in sediments [72]. Other non-algal sources of DMSP have also recently been identified, adding to the overall complexity of this cycle [24,25].
Algal intracellular DMSP is released into the water column by a variety of processes, e.g., exudation [53], cell lysis, or “sloppy” grazing [73]. Very little dissolved DMSP appears to be released by healthy, growing phytoplankton, but stressed or senescent cells and those which are infected by viruses contribute significantly to the dissolved pool [53,54]. The fate of this dissolved DMSP is crucial because either assimilation into bacterial biomass or conversion via demethylation will not lead to DMS production, thus reducing the potential for any climate regulatory effect. Moreover, it is known that only a small fraction of the DMSP that is produced is converted to DMS and thence ventilated to the atmosphere, even though, as already indicated, the contribution of DMS to the overall atmospheric sulfate burden is very significant. This raises the intriguing possibility that even a small change to the balance of these bacterially mediated processes (e.g., due to ocean warming) may cause a significant change to the DMS yield and its flux to the atmosphere.
2.2. Role of Heterotrophic Organisms
The uptake and assimilation of dissolved DMSP can satisfy the energy, carbon, and sulfur demands of entire marine bacterial communities [49]. A case in point is the SAR11 (Pelagibacterales) clade, which is of ancient evolutionary origin and comprises the most abundant and ubiquitous clade of heterotrophic marine bacteria in the oceans, estimated to make up 25% of the plankton community [74]. Tripp et al. [75] observe that assimilatory sulfate reduction is deficient in many SAR11 populations, which suggests that a requirement for exogenous reduced organic sulfur from organosulfur compounds, e.g., DMSP, methionine, and methanethiol, is widespread within the SAR11 clade.
Furthermore, O’Brien et al. [76] point out that members of the marine Roseobacter group (which are common in coastal waters) can establish mutualistic relationships with phytoplankton that are, in part, maintained by exchanges of DMSP. A similar sulfonate-based mutualistic interdependence between phytoplankton and SAR11 organisms has also been found in the North Pacific [77].
From the conceptual model presented in Figure 1, it is evident that the DMS yield will depend on the growth rate and biomass of the heterotrophic bacterial community [49] and on the relative importance of the enzymatic cleavage versus demethylation pathways. The demethylation pathway first converts DMSP to methylmercaptopropionate (MMPA) and subsequently to methanethiol (MeSH) or mercaptopropionate (MPA). The first step of this alternative pathway is crucial to marine sulfur emissions, because it removes a methyl group from DMSP and eliminates DMS as a possible degradation product [78]. The environmental factors that govern the utilization of one pathway over the other, and ultimately the release of DMS to the atmosphere, are uncertain. However, recent work suggests that the concentrations of DMSP that are most relevant for the bacterial production of DMS may not be the levels present in bulk seawater, but instead those existing in microscale hotspots such as near the surface of algal cells [64,79].
Measurements of DMS yield [80,81,82,83] vary widely (1–45%) depending on the region and the seasonal stage of the phytoplankton growth cycle. We note that DMS can also be produced by bacterial reduction of DMSO, the conversion by-product of DMS due to photolytic or bacterial oxidation [84], however the amount of DMS produced via this process does not seem to be significant [85].
Members of the Roseobacter group, a ubiquitous group of marine bacteria, are well-known MeSH producers [86], and in waters rich in these bacteria, consumption of DMSP via demethylation can be an order of magnitude larger than DMS production via cleavage. Kiene et al. [49] posited that bacterioplankton will prefer the demethylation/demethiolation over the lyase pathway at low dissolved DMSP concentrations. This is because this pathway provides more energetic benefits, and it is a relatively economic way to assimilate reduced sulfur. At higher dissolved DMSP concentrations, the DMSP that is not assimilated is then available to the cleavage pathway. It is thus likely that DMS yield through the enzymatic cleavage pathway will increase if bacterial sulfur demand is met with excess sulfur [82], which can occur during the later stages of a phytoplankton bloom [9].
2.3. Impacts of Changes to Ocean Temperature and pCO2
The global oceans face multiple CO2-driven impacts in the coming decades, including ocean acidification (OA), warming, deoxygenation, and loss of sea ice cover [87], all of which will likely affect the carbon budget of the biosphere which depends largely on the balance between the uptake of carbon by phytoplankton photosynthesis and its remineralization by heterotrophs [88]. Warming of the ocean will directly and indirectly impact the growth rates of phytoplankton, potentially altering global primary production and leading to major shifts in the functional trait composition of marine phytoplankton communities [89,90]. The directionality of the productivity response, either positive or negative, will depend on the temperature sensitivity of phytoplankton both within and across species [90], with higher temperatures often leading to a shift towards smaller species such as cyanobacteria [91,92,93]. Similarly, changes in temperature have been found to regulate the metabolic rates of bacteria in marine environments [94]. Thus, the balance between autotrophic production and heterotrophic respiration is profoundly affected by the environmental temperature [95]. However, attempting to describe the response of bacteria to temperature is challenging since other environmental factors that covary with temperature, such as nutrient concentration or primary productivity, may themselves have a larger effect on bacterial properties [96]. Despite the complex interactions between temperature, substrate availability, cell size, and abundance in the ocean, strong apparent increases with temperature in the abundance of picoplankton (<2 μm in diameter), which includes all free-living heterotrophic bacteria, have consistently been reported across different aquatic ecosystems [97]. However, the combined impacts of temperature and pCO2 increases appear to confound the situation, with decreases in phytoplankton biomass combined with increases in bacterial production reported in the Arctic [98].
Over the last two decades, numerous studies have explored the effect of climate change on the broad structure and functioning of the marine food web and the possible changes in net primary production (NPP). This has been attempted through both modelling and mesocosm experiments [99,100,101,102,103,104]. However, owing to computational constraints, temperature sensitivities of recycling processes and especially the microbial loop are ignored in most global models [104,105], notwithstanding that it has long been recognized that bacteria dominate the abundance, diversity, and metabolic activity of the ocean [106] and that heterotrophic processes such as bacterial degradation are known to be temperature-dependent [107]. Brewer and Peltzer [108] provide an excellent review of the broader aspects of this issue in the context of ocean de-oxygenation and bemoan the general neglect of the temperature dependence of microbial decomposition rates in the literature.
2.4. Modeling
Prognostic DMS models have been under development since the early 1990s [109]. Most of the initial efforts were built on food web compartment model approaches [110], and focused on understanding the relationship between phytoplankton bloom dynamics and DMS (P) production in specific regions, e.g., the Arctic [111,112], the Southern Ocean [113], the North Atlantic [114], and the Northwest Pacific [115]. Most of these modeling attempts have been to some degree constrained by limited data on the model parameters. Although sensitivity analyses and other techniques can assist with model calibration at the local scale [116], a major problem in applying such models at a global scale is the site-specific nature of parameter calibration in the field.
The difficulty of applying DMS prognostic models at the global scale has prompted efforts to develop globally applicable diagnostic schemes that predict DMS seawater concentration based on variables such as surface chlorophyll and light, which can be retrieved using satellite remote sensing algorithms [62,117,118,119,120]. These schemes are based on linear regression techniques to estimate DMS concentrations using one or more predictors. Some schemes have demonstrated reasonably good performance at the global scale, but their predictive power is generally less at the regional scale [121]. Currently, none of the global schemes have been accepted universally, with the main deficiencies related to regional biases in remotely sensed chlorophyll (which cause underestimation of DMS in the Southern Ocean) and the inability to reproduce observed variability in “DMS hotspots” such as the northeast sub-Arctic Pacific [122].
Notwithstanding the lack of a globally applicable DMS prediction algorithm [123], there have been numerous attempts to simulate the impact of climate change on future DMS seawater concentration and flux to the atmosphere [37,41,42,124,125,126,127]. Typically, these studies have coupled a global climate model with either a biogeochemical DMS model or empirical prediction algorithm, where the relevant future forcings such as sea surface temperature (SST), surface wind speed, and ocean mixed layer depth drive the future change in DMS seawater concentration.
Bock et al. [39] examine trends of surface ocean DMS concentration and flux of four Earth system models (ESMs: CNRM-ESM2-1, MIROC-ES2L, NorESM2-LM, and UKESM1-0-LL) over the recent past (1980–2009) and into the future, using Coupled Model Intercomparison Project 6 (CMIP6) simulations. The four ESMs disagree on the sign of the trend of the global DMS flux from 1980 onwards, with two models showing an increase and two models a decrease. At the global scale, these trends are dominated by changes in surface DMS concentrations in all models, irrespective of the air–sea flux parameterization used. Three of the models consistently show that changes in DMS concentrations are correlated with changes in marine productivity; however, marine productivity is poorly constrained in the current generation of ESMs. In contrast, a consensus is found among all models at polar latitudes, where an increasing trend is predominantly driven by the simulated retreat in sea-ice extent, however, the magnitude of this trend between models differs by a factor of three. Given the variety of biogeochemical model formulations used, it is not surprising that the simulated changes in DMS under warming vary significantly. In summary, there is no agreement in the literature with regards to the sign and amplitude of the trend in DMS concentration and flux in the future [128].
As indicated above, it is likely that the simulated variability in future DMS trends is related to the inability of current ESMs to accurately project future changes in marine primary production. Depending on whether the ESM formulation has biological processes that are temperature-dependent or not, simulated NPP can either increase or decrease under projected climate change [129]. Indeed, a comparison of nine marine ecosystem model projections of changes in global marine NPP showed quite large uncertainties, especially at low latitudes [130]. Most global ESMs used for carbon sink projections in the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC AR5) ignored the impact of warming on organic carbon remineralization and the biological carbon pump [105]. Segschneider and Bendtsen [104] quantified the impact of including temperature-dependent remineralization (TDR), modifying the CMIP5 model MPI-ESM and its marine biogeochemistry model HAMOCC5.2, and projected an ~0.18 PgC year−1 reduction in ocean carbon uptake by 2100 under the high-emission scenario RCP8.5. The inclusion of more complex ecological parametrizations in the ESM simulates a more modest decline in the ocean carbon sink capacity of ~0.06 PgC year−1 during the 21st century [105], compared to the estimated 2020 ocean carbon sink uptake rate of 3.0 ± 0.4 PgC year−1 [131].
2.5. Laboratory Experiments
Over the past decade, several mesocosm and ship-board bioassay experiments have examined the impact of future increased temperature and pCO2 level on the DMS (P) cycle by perturbing either one or both variables simultaneously. Avgoustidi et al. [132] looked at the change from an approximate doubling of pCO2 (700 ppm) in Emiliania huxleyi and found that DMS production was roughly halved, which they attributed to a change in intracellular DMSP at lower pH values. Spielmeyer and Pohnert [133] analyzed DMSP in separate cultures of diatoms (Thalassiosira pseudonana, Phaeodactylum tricornutum) and a prymnesiophyte (Emiliania huxleyi) under the influence of increased temperature, by 6 °C, and elevated CO2 to 790 ppmv. The diatoms and prymnesiophytes revealed opposite trends for DMSP. In diatoms, increased CO2 and temperature led to decreased DMSP concentrations, however elevated levels of this metabolite under the influence of these parameters were observed for Emiliania huxleyi.
Arnold et al. [134] also report measurements of DMSP and DMS concentrations in pH-stated cultures of Emiliania huxleyi. Four different environmental conditions were tested: ambient, elevated CO2 (+CO2), elevated temperature (+T), and elevated temperature and CO2 (+TCO2). In comparison to the ambient treatment, average DMS production was about 50% lower in the +CO2 treatment. Importantly, temperature had a strong effect on DMS production, and the impacts outweighed the effects of a decrease in pH, with a more than doubling of intracellular DMSP.
Park et al. [135] conducted a series of perturbation experiments in Korean coastal waters and found that at both ambient temperature and ~+2 °C warmer, an increase in pCO2 (160–830 ppm) favored the growth of large diatoms, which outcompeted other phytoplankton species and reduced the growth rate of smaller, DMSP-rich dinoflagellates. This decreased the grazing rate of heterotrophic dinoflagellates, resulting in reduced DMS production via grazing activity, and highlights the influence of community composition changes on future DMS production under warming.
Hopkins and Archer [136] conducted 96 h bioassay ship-board pCO2 perturbation experiments in NW European waters and found that the changes induced consistent, marked increases in DMS and decreases in DMSP, in contrast to results from longer-term mesocosm experiments. Bacterially mediated DMS processes appeared to be insensitive to ocean acidification, with no detectable effects on dark rates of DMS consumption and gross production and no consistent response seen in bacterial abundance [136].
Li et al. [137] conducted a similar experiment with a culture of the dinoflagellate Amphidinium carterae to investigate the effects of elevated CO2 concentration and temperature on the growth and production of DMSP and DMS, however they found no significant effects on the concentrations and cell-normalized concentrations of DMSP or DMS. Saint-Macary et al. [138] observed a reduction in DMSP at warmer temperatures which was associated with changes in the phytoplankton community, and in particular with small flagellate biomass. A smaller decrease in DMS concentration was measured in the treatments relative to other studies.
Significant positive correlations were found between bacterial production and concentrations of DMS in Canadian waters during autumn. The 13-day mesocosm experiments were characterized by blooms of diatoms, with the results pointing to temperature-associated enhancement of bacterial DMSP metabolism and large increases to DMS (>200%), negating any decreases due to acidification [139]. However, later work by the same group of authors in the same Canadian waters during summer found that doubling and tripling the pCO2 resulted in a 15% and 40% decline in average concentrations of DMS compared to the control. Results from S-35-DMSPd uptake assays indicated that neither concentrations nor microbial scavenging efficiency of dissolved DMSP was affected by increased pCO2. However, their results show a reduction of the mean microbial yield of DMS by 34% and 61% in the 2 × pCO2 and 3 × pCO2 treatments, respectively [140].
Clearly, the results of these perturbation experiments are varied and appear to sensitively depend on the phytoplankton groups present, the ocean region examined (polar or temperate), and the perturbation applied. Subtle interactions are also evident, which can lead to counter-intuitive results. For instance, in studies of natural communities that are dominated by the DMSP-rich species Emiliania huxleyi, it would seem likely that the adverse effect of increased pH on the growth of this calcifying organism would also result in a negative impact on the overall production of both DMSP and DMS [141], but this is not always the case, as indicated by the results of Arnold et al. [134].
The range of different results in these experiments makes it difficult to present a synthetic view of the impact of temperature and/or pCO2 changes on future DMS (P) production. It is apparent that the results are sensitive not only to the phytoplankton functional group present but also to the length and type of experiment (mesocosm versus bioassay) and thus the potential for shifts in the community composition to occur.
3. Projected Ocean Warming and Acidification
3.1. Global CMIP6 Climate Projections
The Coupled Model Intercomparison Project Phase 6 (CMIP6) provides climate projections for a range of scenarios to understand the past, present, and future climate. For a better understanding of the impact of the projected climate change on SST, mixed layer depth (MLD), and pH, these are examined for both a contemporary (2001–2020) and end of century (2081–2100) climate, as simulated by CMIP6 models under a SSP2-4.5 and SSP5-8.5 Shared Socioeconomic Pathway (SSP). The possible impact of ocean warming and acidification on DMS (P) production and cycling is then discussed below.
Model output for monthly mean SST (°C) and MLD (m) was obtained from the Australian Community Climate and Earth-System Simulator-Coupled Model version 2 (ACCESS-CM2) [142] for the CMIP6 historical, SSP2-4.5, and SSP5-8.5 experiments. Monthly mean pH was also obtained from the Geophysical Fluid Dynamics Laboratory Earth System Model version 4 (GFLD-ESM4) [143].
The historical simulations run to 2014, with solar variability, volcanic aerosols, and anthropogenic-driven changes in atmospheric composition (GHG and aerosols) forced by datasets that are largely based on observations [144]. The contemporary climatology was extended from 2014 to 2020 by including an average of the CMIP6 model output for the SSP2-4.5 and SSP5-8.5 scenario experiments. For the future SSP scenarios, variables are simulated from 2015 onwards under the respective emissions trajectory. For SSP2-4.5, a medium positive radiative forcing (+4.5 Wm−2) is expected by 2100, assuming a shift towards renewable energy and sustainable development [145]. For SSP5-8.5, a high positive radiative forcing (+8.5 Wm−2) is expected by 2100, assuming that development will be largely driven by fossil fuels [146]. Data were obtained for the r1i1p1f1 ensemble and are available from the Earth System Grid Federation (https://esfg-node.llnl.gov/search/cmip6 (accessed on 19 June 2022)).
3.1.1. Ocean Warming
Ocean warming poses a serious threat to many marine ecosystems. Anthropogenic GHG emissions trap solar radiation within the Earth’s atmosphere, leading to an accumulation of excess heat energy. More than 90% of this excess heat is stored in the ocean, leading to a 0.5 °C rise in global mean SST over the past 40 years [147]. By the end of this century, global mean SST is predicted to increase by a further 1.8 and 3.2 °C under a SSP2-4.5 and SSP5-8.5 climate, respectively (Figure 2).
Temperature impacts the physiology of marine organisms, ocean currents, mixing, and the availability of nutrients in the ocean. Rising SST is predicted to contribute to a stratification of the upper ocean layers and an overall reduction in global mixed layer depth, particularly in the Southern Hemisphere mid–high latitudes and North Atlantic Ocean, where present-day MLD is deepest (Figure 3). Reduced vertical mixing in a more stratified ocean will likely have impacts on the marine sulfur cycle by reducing nutrient concentrations and limiting productivity in the photic zone.
3.1.2. Ocean Acidification
Ocean acidification occurs due to increased absorption of atmospheric CO2. Dissolution of CO2 forms carbonic acid, which readily dissociates into bicarbonate ions and protons, thereby reducing pH. Current anthropogenic CO2 emissions are 10.1 ± 0.5 Gt C year−1 [148]. Approximately 30% of anthropogenic CO2 is absorbed by the ocean and has led to a decrease in global mean ocean pH of 0.1 units over the past 100 years [147]. Despite a reduction in CO2 emissions by the end of this century under an SSP2-4.5 scenario, global mean ocean pH is predicted to decline by −0.18 (See Figure 4). The decline in pH is more extreme under the SSP5-8.5 scenario (−0.36), where little or no effort is made to constrain anthropogenic emissions [146]. Ocean acidification is already impacting on the physiology of marine species, particularly those which are dependent on bioavailable carbonate such as coral reefs.
3.2. Impacts on DMS Cycling in the Polar Oceans
The polar oceans contribute a significant proportion of global DMS emissions and are warming more rapidly than lower latitudes. Factors such as rapid sea ice retreat in the Arctic [149] and the presence of DMSP-rich species such as Phaeocystis antarctica in the Southern Ocean [150] contribute to the potential for major changes in the structure of polar ecosystems and DMS production as warming unfolds [151]. The role of heterotrophic bacteria in the polar regions has been unclear as it was thought that they would be less active because of low temperatures, however Kirchman et al. [152] suggest that the lower activity is partly temperature-driven but mainly due to lower dissolved organic matter (DOM) inputs, a situation which could be exacerbated by increased stratification under warming. In a decade-long study at a highly productive site (Palmer Station) in the Western Antarctic Peninsula (WAP) site, Kim and Ducklow [153] report that bacterial production (BP) was only 4% of the primary production (PP), consistent with the low BP:PP ratios observed in other studies of polar waters. The degree of bottom-up control on bacterial abundance was moderate and relatively consistent across entire growing seasons, suggesting that bacteria in the coastal WAP are under DOC limitation. Temperature also influenced BP rates, though its effect was weaker than the supply of DOC, which is consistent with the findings of Kirchman et al. [152].
The polar oceans are characterized by high dissolved inorganic carbon (DIC) concentrations and a low carbonate system buffering capacity, mainly due to the increased solubility of CO2 in cold waters [154]. This makes these polar regions particularly susceptible to the impacts of OA [155], although there is also evidence of regional variability between the Arctic and Southern Ocean due to differing micro-nutrient regimes [156]. Although quite limited, analyses of the effect of OA on DMS production suggest that OA could significantly decrease the algal biomass and inhibit DMS production during the seasonal phytoplankton bloom in the Arctic [157,158].
In contrast, Hopkins et al. [159] report findings from seven summertime ship-board microcosm experiments undertaken in a variety of locations in the Arctic Ocean and Southern Ocean, which reveal no significant effects of short-term OA on the net production of DMS by planktonic communities. In a follow-up meta-analysis, Hopkins et al. [160] examined experiments in both temperate and polar waters and found clear regional differences in the DMS response to OA, leading them to conclude that future temperate oceans could be more sensitive to OA, while perhaps surprisingly, DMS emissions from the polar oceans may remain relatively unchanged.
3.3. Impacts on the Tropical Ocean and Coral Reefs
Ocean warming and acidification pose serious threats to tropical coral reef ecosystems, with impacts already evident through more frequent and severe coral bleaching events [161,162,163], reduced coral cover and rates of larval recruitment [164], and a decline in coral calcification and growth rates [165].
Reef-building Scleractinian corals have relatively narrow temperature tolerance ranges [163,166,167], and many already live in regions where SST is close to their thermal tolerance thresholds, meaning that increases in SST as small as 0.5 °C could be sufficient to cause coral bleaching [168]. It is predicted that annual severe coral bleaching events will affect more than 95% of tropical coral reefs by 2050 [169]. At 1.5 °C above pre-industrial SST, coral thermal refugia zones and recovery periods are predicted to decline [170,171] and ongoing coral reef degradation may become unavoidable. By the end of this century, CMIP6 models predict tropical SST to increase by a respective 2.0 and 3.6 °C under an SSP2-4.5 and SSP5-8.5 scenario (Figure 5a), with likely detrimental impacts on the coral reef ecosystem structure. Mixed layer depths, which are already shallow in the tropical ocean, shoal further under warming scenarios (Figure 5b).
Coral reef ecosystems are important regional sources of dissolved and atmospheric sulfur, emitting an estimated 0.05–0.08 Tg year−1 of DMS (equivalent to 0.025–0.04 Tg year−1 S) [172]. Reported concentrations of atmospheric DMS (DMSa) in the Great Barrier Reef (GBR), Australia, average approximately 30 ppt in winter and 100 ppt in summer [173,174], and can exceed 500 ppt over coral reefs that have been exposed to the atmosphere during low tide [174,175]. On one occasion at Heron Island in the southern GBR, DMSa concentration reached 1149 ppt when the coral reef flat was thermally and osmotically shocked by rainfall [174]. These concentrations are up to two orders of magnitude higher than the background ocean, meaning that coral reefs are important hotspots of marine sulfur. However, the future contribution of coral reefs to the dissolved and atmospheric sulfur pool will depend on the ability of corals to acclimate to their rapidly changing environment.
DMSP biosynthesis and cleavage to DMS is upregulated in corals in response to thermal and irradiance stress [175,176,177]. When high temperatures combine with elevated light levels, the photosystems of zooxanthellae and free-living algae can become photo-inhibited and produce ROS [18,178,179,180]. If conditions persist, ROS accumulate and cause oxidative damage in the coral holobiont [167,180,181]. To cope with oxidative stress, corals may change the composition of their microbiome (e.g., zooxanthellae switching or shuffling) to enhance temperature tolerance [182] or may expel zooxanthellae and become bleached [167,181,183]. DMS (P) plays an important role in mitigating oxidative stress in corals [178] and marine algae [18] by scavenging ROS and forming DMSO.
Shifts in the coral holobiont microbiome can result in a change in DMS (P) production [184,185]. DMS (P) concentrations vary between endosymbiont types, with more temperature-sensitive types (e.g., clade C) typically containing higher concentrations than temperature-tolerant types (e.g., clade D) when exposed to elevated temperatures [182,186]. However, tolerance thresholds vary between coral and endosymbiont species [187], and other factors such as assemblage complexity [19] can influence DMS (P) production in coral reefs.
If corals can withstand rising ocean temperatures through natural means, such as the recruitment of temperature-tolerant endosymbionts [182,184], DMS production and emissions from coral reefs may increase in the future. However, observational studies have shown that seawater surface and atmospheric DMS concentrations in the GBR increase with SST, until SST approaches coral thermal tolerance thresholds [175,188]. Beyond this threshold, ambient DMS concentrations can decline [175,185,189], possibly due to an upregulation of the coral and algal antioxidant response discussed above. Therefore, if corals cannot acclimate, DMS concentrations and emissions may decline due to an upregulation of the coral antioxidant response [178], or due to increased coral bleaching and mortality leading to reduced coral cover [164].
Exacerbating the impacts of ocean warming, ocean acidification has already led to a decline in the bioavailability of carbonate ions [190], limiting calcification and growth of calcareous organisms. In the GBR, skeletal density and linear extension rates of massive Porites corals have declined by 0.36% and 1.02%, respectively [165]. Although the impacts of ocean acidification on the coral reef sulfur cycle are unclear, a decline in coral cover will likely shift the balance of coral–algal dynamics in reef ecosystems.
As for ocean regions, algal and microbial communities play an important role in the production and cycling of DMS (P) in the coral holobiont and in coral reef waters [177]. Increasing SST and enhanced stratification (Figure 5) may limit nutrient availability and phytoplankton productivity in the GBR. However, GBR waters are predicted to become more eutrophic with increasing inputs of nitrogen and phosphorus from agricultural runoff and soil erosion [191,192,193,194]. Improving water quality is a key commitment of the Australian Government’s Reef 2050 Long-Term Sustainability Plan.
Further, corals and algae compete for light, nutrients, and space in the reef matrix. Predicted declines in coral cover due to ocean warming, acidification, predation, and declining water quality may lead to an increase in algal biomass in coral reefs. Increased algal biomass is often observed after mass coral bleaching and mortality events [195], impeding coral growth and reducing opportunities for new coral establishment [196]. An increase in alternative sulfur sources (e.g., through riverine inputs) reduces sulfur limitations and thus the need for microbial DMSP demethylation, thereby increasing DMS yields via the alternative cleavage pathway [22].
4. Conclusions
Ecological interactions between marine bacteria and phytoplankton are now recognized as playing a central role in the ocean’s major biogeochemical cycles. How these interactions may be modified as a result of climate change is the focus of this review, which examined the role of heterotrophic bacterioplankton in the cycling of a suite of organo-sulfur compounds, such as DMS. This volatile compound and its precursor DMSP are ubiquitous in the oceans, being synthesized by a large range of marine organisms from algae to corals. After ventilation, DMS constitutes the largest source of natural sulfur to the atmosphere and is thus also an important contributor to the atmospheric burden of sulfate aerosol with key relevance to the Earth’s radiative balance and climate.
Due to its synthesis by numerous marine organisms, DMSP and its related compounds are an important source of DOM for bacterial heterotrophy in the surface ocean, where it has been estimated to contribute up to 100% of the sulfur requirements of some important marine bacteria groups. There is growing evidence that sulfonates serve as an ecologically important currency for nutrient and energy exchange between autotrophs and heterotrophs, highlighting the importance of organo-sulfates in regulating ecosystem function. This suggests that climate change impacts on the synthesis and cycling of DMS (P) could have important future consequences for the structure of the marine food web.
However, perturbation experiments seeking to understand the impact of warming and OA on the synthesis of DMS have so far yielded equivocal results. The range of different results makes it difficult to provide a synthetic view of the impact of temperature and/or pCO2 changes on future DMS (P) production. It is apparent that the results are sensitive not only to the phytoplankton functional group present but also to the length and type of experiment conducted.
Although numerous attempts have been made to numerically model the impact of warming on the DMS (P) cycle, there is no general agreement in the literature with regards to the sign and amplitude of the future trend in DMS concentration and sea-to-air flux. Importantly, these efforts are also constrained (due to current knowledge of the process and computational limits) by a lack of an explicit temperature-dependent parameterization of the key bacterial-mediated processes, such as demethylation and enzymatic cleavage. Considering the critical roles DMS and its precursor DMSP appear to play in supplying carbon and sulfur to marine heterotrophs, future efforts should focus on informing the many uncertainties still apparent in the literature regarding the ecological role of these compounds, and especially the nexus between future CC impacts on marine bacteria and the possible resultant changes in the marine DMS (P) cycles.
Author Contributions
Conceptualization A.G. and R.J.; Software, R.J.; Writing—original draft preparation, A.G. and R.J.; writing—review and editing, A.G. and R.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The CMIP6 model output used in this analysis [142,143] is available from the Earth System Grid Federation (https://esfg-node.llnl.gov/search/cmip6/ (accessed on 19 June 2022)).
Acknowledgments
The authors acknowledge the world Climate Research Programme’s Working Group on Coupled Modelling, which is responsible for CMIP, and the Commonwealth Scientific and Industrial Research Organisation (CSIRO), Australian Research Council Centre of Excellence for Climate System Science (and the National Oceanic and Atmospheric Administration Geophysical Fluid Dynamics Laboratory for producing and making available their model output. The model output used in this analysis is available from the Earth System Grid Federation (https://esfg-node.llnl.gov/search/cmip6/ (accessed on 19 June 2022)). The U.S. Department of Energy’s Program for Climate Model Diagnosis and Intercomparison provides coordinating support for CMIP and led development of software infrastructure in partnership with the Global Organization for Earth System Science Portals.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Andreae, M.O.; Crutzen, P.J. Atmospheric aerosols: Biogeochemical sources and role in atmospheric chemistry. Science 1997, 276, 1052–1058. [Google Scholar] [CrossRef]
- Shaw, G.E. Bio-controlled thermostasis involving the sulphur cycle. Clim. Chang. 1983, 5, 297–303. [Google Scholar] [CrossRef]
- Charlson, R.J.; Lovelock, J.; Andreae, M.O.; Warren, S.G. Oceanic phytoplankton, atmospheric sulphur, cloud albedo and climate. Nature 1987, 326, 655–661. [Google Scholar] [CrossRef]
- Brévière, E.H.G.; Bakker, D.C.E.; Bange, H.W.; Bates, T.S.; Bell, T.G.; Boyd, P.W.; Duce, R.A.; Garçon, V.; Johnson, M.T.; Law, C.S.; et al. Surface ocean-lower atmosphere study: Scientific synthesis and contribution to Earth system science. Anthropocene 2015, 12, 54–68. [Google Scholar] [CrossRef]
- Ayers, G.P.; Cainey, J.M. The CLAW hypothesis: A review of the major developments. Environ. Chem. 2007, 4, 366–374. [Google Scholar] [CrossRef]
- Liss, P.S.; Marandino, C.A.; Dahl, E.E.; Helmig, D.; Hintsa, E.J.; Hughes, C.; Johnson, M.T.; Moore, R.M.; Plane, J.; Quack, B. Short-lived trace gases in the surface ocean and the atmosphere. In Ocean-Atmosphere Interactions of Gases and Particles; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–54. [Google Scholar]
- Wang, W.L.; Song, G.; Primeau, F.; Saltzman, E.S.; Bell, T.G.; Moore, J.K. Global ocean dimethyl sulfide climatology estimated from observations and an artificial neural network. Biogeosciences 2020, 17, 5335–5354. [Google Scholar] [CrossRef]
- Chin, M.A.; Jacob, D.J. Anthropogenic and natural contributions to tropospheric sulfate: A global model analysis. J. Geophys. Res.-Atmos. 1996, 101, 18691–18699. [Google Scholar] [CrossRef]
- Kilgour, D.B.; Novak, G.A.; Sauer, J.S.; Moore, A.N.; Dinasquet, J.; Amiri, S.; Franklin, E.B.; Mayer, K.; Winter, M.; Morris, C.K. Marine gas-phase sulfur emissions during an induced phytoplankton bloom. Atmos. Chem. Phys. 2022, 22, 1601–1613. [Google Scholar] [CrossRef]
- Smith, S.J.; van Aardenne, J.; Klimont, Z.; Andres, R.J.; Volke, A.; Delgado Arias, S. Anthropogenic sulfur dioxide emissions: 1850–2005. Atmos. Chem. Phys. 2011, 11, 1101–1116. [Google Scholar] [CrossRef]
- Mitchell, J.F.; Johns, T.; Gregory, J.M.; Tett, S. Climate response to increasing levels of greenhouse gases and sulphate aerosols. Nature 1995, 376, 501–504. [Google Scholar] [CrossRef]
- Westervelt, D.M.; Horowitz, L.W.; Naik, V.; Golaz, J.C.; Mauzerall, D.L. Radiative forcing and climate response to projected 21st century aerosol decreases. Atmos. Chem. Phys. 2015, 15, 12681–12703. [Google Scholar] [CrossRef]
- Aas, W.; Mortier, A.; Bowersox, V.; Cherian, R.; Faluvegi, G.; Fagerli, H.; Hand, J.; Klimont, Z.; Galy-Lacaux, C.; Lehmann, C.M.B.; et al. Global and regional trends of atmospheric sulfur. Sci. Rep. 2019, 9, 953. [Google Scholar] [CrossRef]
- Quinn, P.K.; Bates, T.S. The case against climate regulation via oceanic phytoplankton sulphur emissions. Nature 2011, 480, 51–56. [Google Scholar] [CrossRef]
- Novak, G.A.; Fite, C.H.; Holmes, C.D.; Veres, P.R.; Neuman, J.A.; Faloona, I.; Thornton, J.A.; Wolfe, G.M.; Vermeuel, M.P.; Jernigan, C.M.; et al. Rapid cloud removal of dimethyl sulfide oxidation products limits SO2 and cloud condensation nuclei production in the marine atmosphere. Proc. Natl. Acad. Sci. USA 2021, 118, e2110472118. [Google Scholar] [CrossRef]
- Fung, K.M.; Heald, C.L.; Kroll, J.H.; Wang, S.; Jo, D.S.; Gettelman, A.; Lu, Z.; Liu, X.; Zaveri, R.A.; Apel, E.C.; et al. Exploring dimethyl sulfide (DMS) oxidation and implications for global aerosol radiative forcing. Atmos. Chem. Phys. 2022, 22, 1549–1573. [Google Scholar] [CrossRef]
- Keller, M.D.; Bellows, W.K.; Guillard, R.L. Dimethyl Sulfide production in marine phytoplankton. In Biogenic Sulfur in the Environment; Saltzman, E.S., Cooper, W.J., Eds.; American Chemical Society: Washington, DC, USA, 1989. [Google Scholar]
- Sunda, W.; Kieber, D.J.; Kiene, R.P.; Huntsman, S. An antioxidant function for DMSP and DMS in marine algae. Nature 2002, 418, 317–320. [Google Scholar] [CrossRef]
- Guibert, I.; Bourdreux, F.; Bonnard, I.; Pochon, X.; Dubousquet, V.; Raharivelomanana, P.; Berteaux-Lecellier, V.; Lecellier, G. Dimethylsulfoniopropionate concentration in coral reef invertebrates varies according to species assemblages. Sci. Rep. 2020, 10, 9922. [Google Scholar] [CrossRef]
- Reisch, C.R.; Moran, M.A.; Whitman, W.B. Bacterial catabolism of dimethylsulfoniopropionate (DMSP). Front. Microbiol. 2011, 2, 172. [Google Scholar] [CrossRef]
- Gregory, G.J.; Boas, K.E.; Boyd, E.F. The organosulfur compound dimethylsulfoniopropionate (DMSP) is utilized as an osmoprotectant by Vibrio species. Appl. Environ. Microbiol. 2020, 87, e02235-20. [Google Scholar] [CrossRef]
- Fernandez, E.; Ostrowski, M.; Siboni, N.; Seymour, J.R.; Petrou, K. Uptake of Dimethylsulfoniopropionate (DMSP) by Natural Microbial Communities of the Great Barrier Reef (GBR), Australia. Microorganisms 2021, 9, 1891. [Google Scholar] [CrossRef]
- Ainsworth, T.; Krause, L.; Bridge, T.; Torda, G.; Raina, J.-B.; Zakrzewski, M.; Gates, R.D.; Padilla-Gamiño, J.L.; Spalding, H.L.; Smith, C. The coral core microbiome identifies rare bacterial taxa as ubiquitous endosymbionts. ISME J. 2015, 9, 2261–2274. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.L.; Gabric, A.J.; Cropp, R.; Woodhouse, M.T. Dimethylsulfide (DMS), marine biogenic aerosols and the ecophysiology of coral reefs. Biogeosciences 2020, 17, 2181–2204. [Google Scholar] [CrossRef]
- Raina, J.B.; Tapiolas, D.M.; Foret, S.; Lutz, A.; Abrego, D.; Ceh, J.; Seneca, F.O.; Clode, P.L.; Bourne, D.G.; Willis, B.L.; et al. DMSP biosynthesis by an animal and its role in coral thermal stress response. Nature 2013, 502, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Haydon, T.D.; Seymour, J.R.; Suggett, D.J. Soft corals are significant DMSP producers in tropical and temperate reefs. Mar. Biol. 2018, 165, 109. [Google Scholar] [CrossRef]
- Van Alstyne, K.L.; Puglisi, M.P. DMSP in marine macroalgae and macroinvertebrates: Distribution, function, and ecological impacts. Aquat. Sci. 2007, 69, 394–402. [Google Scholar] [CrossRef]
- Sanchez, K.J.; Chen, C.-L.; Russell, L.M.; Betha, R.; Liu, J.; Price, D.J.; Massoli, P.; Ziemba, L.D.; Crosbie, E.C.; Moore, R.H.; et al. Substantial Seasonal Contribution of Observed Biogenic Sulfate Particles to Cloud Condensation Nuclei. Sci. Rep. 2018, 8, 3235. [Google Scholar] [CrossRef]
- Korhonen, H.; Carslaw, K.S.; Spracklen, D.V.; Mann, G.W.; Woodhouse, M.T. Influence of oceanic dimethyl sulfide emissions on cloud condensation nuclei concentrations and seasonality over the remote Southern Hemisphere oceans: A global model study. J. Geophys. Res.-Atmos. 2008, 113, D15204. [Google Scholar] [CrossRef]
- Xu, F.; Yan, S.-B.; Zhang, H.-H.; Wu, Y.-C.; Ma, Q.-Y.; Song, Y.-C.; Zhuang, G.-C.; Yang, G.-P. Occurrence and cycle of dimethyl sulfide in the western Pacific Ocean. Limnol. Oceanogr. 2021, 66, 2868–2884. [Google Scholar] [CrossRef]
- Vallina, S.M.; Simo, R. Strong Relationship Between DMS and the Solar Radiation Dose over the Global Surface Ocean. Science 2007, 315, 506–508. [Google Scholar] [CrossRef]
- Galí, M.; Saló, V.; Almeda, R.; Calbet, A.; Simó, R. Stimulation of gross dimethylsulfide (DMS) production by solar radiation. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Lawrence, M.G. An empirical analysis of the strength of the phytoplankton-dimethylsulfide-cloud-climate feedback cycle. J. Geophys. Res. 1993, 98, 20663–20673. [Google Scholar] [CrossRef]
- Foley, J.A.; Taylor, K.E.; Ghan, S.J. Planktonic dimethylsulfide and cloud albedo: An estimate of the feedback response. Clim. Chang. 1991, 18, 1–15. [Google Scholar] [CrossRef]
- Gabric, A.J.; Whetton, P.H.; Boers, R.; Ayers, G.P. The impact of simulated climate change on the air-sea flux of dimethylsulphide in the subantarctic Southern Ocean. Tellus B 1998, 50, 388–399. [Google Scholar] [CrossRef]
- Six, K.D.; Kloster, S.; Ilyina, T.; Archer, S.D.; Zhang, K.; Maier-Reimer, E. Global warming amplified by reduced sulphur fluxes as a result of ocean acidification. Nat. Clim. Chang. 2013, 3, 975–978. [Google Scholar] [CrossRef]
- Gunson, J.R.; Spall, S.A.; Anderson, T.R.; Jones, A.; Totterdell, I.J.; Woodage, M.J. Climate sensitivity to ocean dimethylsulphide emissions. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef]
- Vallina, S.M.; Simo, R.; Manizza, M. Weak response of oceanic dimethylsulfide to upper mixing shoaling induced by global warming. Proc. Natl. Acad. Sci. USA 2007, 104, 16004–16009. [Google Scholar] [CrossRef]
- Bock, J.; Michou, M.; Nabat, P.; Abe, M.; Mulcahy, J.P.; Olivie, D.J.L.; Schwinger, J.; Suntharalingam, P.; Tjiputra, J.; van Hulten, M.; et al. Evaluation of ocean dimethylsulfide concentration and emission in CMIP6 models. Biogeosciences 2021, 18, 3823–3860. [Google Scholar] [CrossRef]
- Szopa, S.; Naik, V.; Adhikary, B.; Artaxo, P.; Berntsen, T.; Collins, W.D.; Fuzzi, S.; Gallardo, L.; Kiendler-Scharr, A.; Klimont, Z. Short-Lived Climate Forcers. In Proceedings of the AGU Fall Meeting 2021, New Orleans, LA, USA, 3–17 December 2021. [Google Scholar]
- Cameron-Smith, P.; Elliott, S.; Maltrud, M.; Erickson, D.; Wingenter, O. Changes in dimethyl sulfide oceanic distribution due to climate change. Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Gabric, A.J.; Simo, R.; Cropp, R.A.; Hirst, A.; Dachs, J. Modeling estimates of the global emission of dimethylsulfide under enhanced greenhouse conditions. Glob. Biogeochem. Cycle 2004, 18, GB2014. [Google Scholar] [CrossRef]
- Qu, B.; Gabric, A.J.; Jackson, R. Contemporary variability in dimethylsulfide flux in the Barents Sea and simulated change under 4×CO2 climate conditions. J. Mar. Syst. 2021, 220, 103573. [Google Scholar] [CrossRef]
- Qu, B.; Gabric, A.J.; Jackson, R. Simulated perturbation in the sea-to-air flux of dimethylsulfide and the impact on polar climate. J. Oceanol. Limnol. 2021, 39, 110–121. [Google Scholar] [CrossRef]
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.S.; Meyer-Reil, L.-A.; Thingstad, F. The ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Cho, B.C.; Azam, F. Major role of bacteria in biogeochemical fluxes in the ocean’s interior. Nature 1988, 332, 441–443. [Google Scholar] [CrossRef]
- Pomeroy, L.R.; Williams, P.J.l.; Azam, F.; Hobbie, J.E. The microbial loop. Oceanography 2007, 20, 28–33. [Google Scholar] [CrossRef]
- Simó, R. Production of atmospheric sulfur by oceanic plankton: Biogeochemical, ecological and evolutionary links. Trends Ecol. Evol. 2001, 16, 287–294. [Google Scholar] [CrossRef]
- Kiene, R.P.; Linn, L.J.; Bruton, J.A. New and important roles for DMSP in marine microbial communities. J. Sea Res. 2000, 43, 209–224. [Google Scholar] [CrossRef]
- Cirri, E.; Pohnert, G. Algae-bacteria interactions that balance the planktonic microbiome. New Phytol. 2019, 223, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Thume, K.; Gebser, B.; Chen, L.; Meyer, N.; Kieber, D.J.; Pohnert, G. The metabolite dimethylsulfoxonium propionate extends the marine organosulfur cycle. Nature 2018, 563, 412–415. [Google Scholar] [CrossRef]
- Dacey, J.W.H.; Wakeham, S.G. Oceanic dimethylsulfide: Production during zooplankton grazing on phytoplankton. Science 1986, 233, 1314–1316. [Google Scholar] [CrossRef]
- Laroche, D.; Vezina, A.; Levasseur, M.; Gosselin, M.; Stefels, J.; Keller, M.; Matrai, P.; Kwint, R. DMSP synthesis and exudation in phytoplankton: A modeling approach. Mar. Ecol. Prog. Ser. 1999, 180, 37–49. [Google Scholar] [CrossRef]
- Malin, G.; Wilson, W.H.; Bratbak, G.; Liss, P.S.; Mann, N.H. Elevated production of dimethylsulfide resulting from viral infection of cultures of Phaeocystis pouchetii. Limnol. Oceanogr. 1998, 43, 1389–1393. [Google Scholar] [CrossRef]
- Yoch, D.C. Dimethylsulfoniopropionate: Its sources, role in the marine food web, and biological degradation to dimethylsulfide. Appl. Environ. Microbiol. 2002, 68, 5804–5815. [Google Scholar] [CrossRef]
- Groene, T. Biogenic production and consumption of dimethylsulfide (DMS) and dimethylsulfoniopropionate (DMSP) in the marine epipelagic zone: A review. J. Mar. Syst. 1995, 6, 191–209. [Google Scholar] [CrossRef]
- Kiene, R.P.; Bates, T.S. Biological removal of dimethylsulfide from seawater. Nature 1990, 345, 702–705. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef]
- Levine, N.M. Putting the spotlight on organic sulfur. Science 2016, 354, 418–419. [Google Scholar] [CrossRef]
- Moran, M.A.; Durham, B.P. Sulfur metabolites in the pelagic ocean. Nat. Rev. Microbiol. 2019, 17, 665–678. [Google Scholar] [CrossRef]
- Ksionzek, K.B.; Lechtenfeld, O.J.; McCallister, S.L.; Schmitt-Kopplin, P.; Geuer, J.K.; Geibert, W.; Koch, B.P. Dissolved organic sulfur in the ocean: Biogeochemistry of a petagram inventory. Science 2016, 354, 456–459. [Google Scholar] [CrossRef]
- Galí, M.; Devred, E.; Levasseur, M.; Royer, S.-J.; Babin, M. A remote sensing algorithm for planktonic dimethylsulfoniopropionate (DMSP) and an analysis of global patterns. Remote Sens. Environ. 2015, 171, 171–184. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef]
- Gao, C.; Fernandez, V.I.; Lee, K.S.; Fenizia, S.; Pohnert, G.; Seymour, J.R.; Raina, J.-B.; Stocker, R. Single-cell bacterial transcription measurements reveal the importance of dimethylsulfoniopropionate (DMSP) hotspots in ocean sulfur cycling. Nat. Commun. 2020, 11, 1942. [Google Scholar] [CrossRef]
- Reisch, C.R.; Moran, M.A.; Whitman, W.B. Dimethylsulfoniopropionate-Dependent Demethylase (DmdA) from Pelagibacter ubique and Silicibacter pomeroyi. J. Bacteriol. 2008, 190, 8018–8024. [Google Scholar] [CrossRef]
- Kiene, R.P.; Linn, L.J. Distribution and turnover of dissolved DMSP and its relationship with bacterial production and dimethylsulfide in the Gulf of Mexico. Limnol. Oceanogr. 2000, 45, 849–861. [Google Scholar] [CrossRef]
- Kiene, R.P.; Nowinski, B.; Esson, K.; Preston, C.; Marin III, R.; Birch, J.; Scholin, C.; Ryan, J.; Moran, M.A. Unprecedented DMSP concentrations in a massive dinoflagellate bloom in Monterey Bay, CA. Geophys. Res. Lett. 2019, 46, 12279–12288. [Google Scholar] [CrossRef]
- Stefels, J.; Steinke, M.; Turner, S.; Malin, G.; Belviso, S. Environmental constraints on the production and removal of the climatically active gas dimethylsulphide (DMS) and implications for ecosystem modelling. Biogeochemistry 2007, 83, 245–275. [Google Scholar] [CrossRef]
- Novak, G.A.; Kilgour, D.B.; Jernigan, C.M.; Vermeuel, M.P.; Bertram, T.H. Oceanic emissions of dimethyl sulfide and methanethiol and their contribution to sulfur dioxide production in the marine atmosphere. Atmos. Chem. Phys. 2022, 22, 6309–6325. [Google Scholar] [CrossRef]
- Li, C.; Yang, G.-P.; Kieber, D.J.; Motard-Côté, J.; Kiene, R.P. Assessment of DMSP turnover reveals a non-bioavailable pool of dissolved DMSP in coastal waters of the Gulf of Mexico. Environ. Chem. 2016, 13, 266–279. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Liu, J.; Zhong, H.; Williams, B.T.; Zheng, Y.; Curson, A.R.J.; Sun, C.; Sun, H.; Song, D.; et al. Bacterial Dimethylsulfoniopropionate Biosynthesis in the East China Sea. Microorganisms 2021, 9, 657. [Google Scholar] [CrossRef]
- Zhuang, G.-C.; Lin, Y.-S.; Bowles, M.W.; Heuer, V.B.; Lever, M.A.; Elvert, M.; Hinrichs, K.-U. Distribution and isotopic composition of trimethylamine, dimethylsulfide and dimethylsulfoniopropionate in marine sediments. Mar. Chem. 2017, 196, 35–46. [Google Scholar] [CrossRef]
- Kasamatsu, N.; Kawaguchi, S.; Watanabe, S.; Odate, T.; Fukuchi, M. Possible impacts of zooplankton grazing on dimethylsulfide production in the Antartic Ocean. Can. J. Fish. Aquat. Sci. 2011, 61, 736–743. [Google Scholar] [CrossRef]
- Giovannoni, S.J. SAR11 Bacteria: The Most Abundant Plankton in the Oceans. Annu. Rev. Mar. Sci. 2017, 9, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Tripp, H.J.; Kitner, J.B.; Schwalbach, M.S.; Dacey, J.W.H.; Wilhelm, L.J.; Giovannoni, S.J. SAR11 marine bacteria require exogenous reduced sulphur for growth. Nature 2008, 452, 741–744. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; McParland, E.L.; Bramucci, A.R.; Siboni, N.; Ostrowski, M.; Kahlke, T.; Levine, N.M.; Brown, M.V.; van de Kamp, J.; Bodrossy, L.; et al. Biogeographical and seasonal dynamics of the marine Roseobacter community and ecological links to DMSP-producing phytoplankton. ISME Commun. 2022, 2, 16. [Google Scholar] [CrossRef]
- Durham, B.P.; Boysen, A.K.; Carlson, L.T.; Groussman, R.D.; Heal, K.R.; Cain, K.R.; Morales, R.L.; Coesel, S.N.; Morris, R.M.; Ingalls, A.E.; et al. Sulfonate-based networks between eukaryotic phytoplankton and heterotrophic bacteria in the surface ocean. Nat. Microbiol. 2019, 4, 1706–1715. [Google Scholar] [CrossRef]
- Howard, E.C.; Henriksen, J.R.; Buchan, A.; Reisch, C.R.; Bürgmann, H.; Welsh, R.; Ye, W.; González, J.M.; Mace, K.; Joye, S.B.; et al. Bacterial Taxa That Limit Sulfur Flux from the Ocean. Science 2006, 314, 649–652. [Google Scholar] [CrossRef]
- Seymour, J.R.; Amin, S.A.; Raina, J.-B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton–bacteria relationships. Nat. Microbiol. 2017, 2, 17065. [Google Scholar] [CrossRef]
- Vila-Costa, M.; Rinta-Kanto, J.M.; Poretsky, R.S.; Sun, S.; Kiene, R.P.; Moran, M.A. Microbial controls on DMSP degradation and DMS formation in the Sargasso Sea. Biogeochemistry 2014, 120, 295–305. [Google Scholar] [CrossRef]
- Motard-Côté, J.; Levasseur, M.; Scarratt, M.; Michaud, S.; Gratton, Y.; Rivkin, R.B.; Keats, K.; Gosselin, M.; Tremblay, J.É.; Kiene, R.P. Distribution and metabolism of dimethylsulfoniopropionate (DMSP) and phylogenetic affiliation of DMSP-assimilating bacteria in northern Baffin Bay/Lancaster Sound. J. Geophys. Res. Ocean. 2012, 117. [Google Scholar] [CrossRef]
- Lizotte, M.; Levasseur, M.; Law, C.S.; Walker, C.F.; Safi, K.A.; Marriner, A.; Kiene, R.P. Dimethylsulfoniopropionate (DMSP) and dimethyl sulfide (DMS) cycling across contrasting biological hotspots of the New Zealand subtropical front. Ocean Sci. 2017, 13, 961–982. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, S.-H.; Tian, J.-Y.; Zhang, Z.-Y.; Zhao, L.-J.; Xu, R.; Yang, G.-P.; Lai, J.-G.; Wang, X.-D. Distribution and Dimethylsulfoniopropionate Degradation of Dimethylsulfoniopropionate-Consuming Bacteria in the Yellow Sea and East China Sea. J. Geophys. Res. Ocean. 2021, 126, e2021JC017679. [Google Scholar] [CrossRef]
- Spiese, C.E.; Kieber, D.J.; Nomura, C.T.; Kiene, R.P. Reduction of dimethylsulfoxide to dimethylsulfide by marine phytoplankton. Limnol. Oceanogr. 2009, 54, 560–570. [Google Scholar] [CrossRef]
- Dixon, J.L.; Hopkins, F.E.; Stephens, J.A.; Schäfer, H. Seasonal Changes in Microbial Dissolved Organic Sulfur Transformations in Coastal Waters. Microorganisms 2020, 8, 337. [Google Scholar] [CrossRef]
- Zubkov, M.V.; Fuchs, B.M.; Archer, S.D.; Kiene, R.P.; Amann, R.; Burkill, P.H. Rapid turnover of dissolved DMS and DMSP by defined bacterioplankton communities in the stratified euphotic zone of the North Sea. Deep-Sea Res. Part II-Top. Stud. Oceanogr. 2002, 49, 3017–3038. [Google Scholar] [CrossRef]
- Gattuso, J.-P.; Magnan, A.; Billé, R.; Cheung, W.W.L.; Howes, E.L.; Joos, F.; Allemand, D.; Bopp, L.; Cooley, S.R.; Eakin, C.M.; et al. Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 2015, 349, aac4722. [Google Scholar] [CrossRef]
- Del Giorgio, P.A.; Duarte, C.M. Respiration in the open ocean. Nature 2002, 420, 379–384. [Google Scholar] [CrossRef]
- Dutkiewicz, S.; Scott, J.R.; Follows, M.J. Winners and losers: Ecological and biogeochemical changes in a warming ocean. Glob. Biogeochem. Cycle 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Barton, S.; Yvon-Durocher, G. Quantifying the temperature dependence of growth rate in marine phytoplankton within and across species. Limnol. Oceanogr. 2019, 64, 2081–2091. [Google Scholar] [CrossRef]
- Schabhüttl, S.; Hingsamer, P.; Weigelhofer, G.; Hein, T.; Weigert, A.; Striebel, M. Temperature and species richness effects in phytoplankton communities. Oecologia 2013, 171, 527–536. [Google Scholar] [CrossRef]
- Ullah, H.; Nagelkerken, I.; Goldenberg, S.U.; Fordham, D.A. Climate change could drive marine food web collapse through altered trophic flows and cyanobacterial proliferation. PLoS Biol. 2018, 16, e2003446. [Google Scholar] [CrossRef]
- Anderson, S.I.; Barton, A.D.; Clayton, S.; Dutkiewicz, S.; Rynearson, T.A. Marine phytoplankton functional types exhibit diverse responses to thermal change. Nat. Commun. 2021, 12, 6413. [Google Scholar] [CrossRef]
- Rivkin, R.B.; Legendre, L. Biogenic carbon cycling in the upper ocean: Effects of microbial respiration. Science 2001, 291, 2398–2400. [Google Scholar] [CrossRef]
- López-Urrutia, Á.; Martin, E.S.; Harris, R.P.; Irigoien, X. Scaling the metabolic balance of the oceans. Proc. Natl. Acad. Sci. USA 2006, 103, 8739–8744. [Google Scholar] [CrossRef]
- Lomas, M.W.; Glibert, P.M.; Shiah, F.K.; Smith, E.M. Microbial processes and temperature in Chesapeake Bay: Current relationships and potential impacts of regional warming. Glob. Chang. Biol. 2002, 8, 51–70. [Google Scholar] [CrossRef]
- Ducklow, H.W.; Morán, X.A.G.; Murray, A.E. Bacteria in the greenhouse: Marine microbes and climate change. In Environmental Microbiology; Mitchell, R., Ji-Dong Gu, J.-D., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2010; pp. 1–31. [Google Scholar]
- Vaqué, D.; Lara, E.; Arrieta, J.M.; Holding, J.; Sà, E.L.; Hendriks, I.E.; Coello-Camba, A.; Alvarez, M.; Agustí, S.; Wassmann, P.F.; et al. Warming and CO2 Enhance Arctic Heterotrophic Microbial Activity. Front. Microbiol. 2019, 10, 494. [Google Scholar] [CrossRef]
- Bopp, L.; Resplandy, L.; Orr, J.C.; Doney, S.C.; Dunne, J.P.; Gehlen, M.; Halloran, P.; Heinze, C.; Ilyina, T.; Seferian, R.; et al. Multiple stressors of ocean ecosystems in the 21st century: Projections with CMIP5 models. Biogeosciences 2013, 10, 6225–6245. [Google Scholar] [CrossRef]
- Lewandowska, A.M.; Boyce, D.G.; Hofmann, M.; Matthiessen, B.; Sommer, U.; Worm, B. Effects of sea surface warming on marine plankton. Ecol. Lett. 2014, 17, 614–623. [Google Scholar] [CrossRef]
- Wohlers, J.; Engel, A.; Zollner, E.; Breithaupt, P.; Jurgens, K.; Hoppe, H.G.; Sommer, U.; Riebesell, U. Changes in biogenic carbon flow in response to sea surface warming. Proc. Natl. Acad. Sci. USA 2009, 106, 7067–7072. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Goldenberg, S.U.; Ferreira, C.M.; Ullah, H.; Connell, S.D. Trophic pyramids reorganize when food web architecture fails to adjust to ocean change. Science 2020, 369, 829–832. [Google Scholar] [CrossRef]
- Sarmento, H.; Montoya, J.M.; Vázquez-Domínguez, E.; Vaqué, D.; Gasol, J.M. Warming effects on marine microbial food web processes: How far can we go when it comes to predictions? Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2137–2149. [Google Scholar] [CrossRef]
- Segschneider, J.; Bendtsen, J. Temperature-dependent remineralization in a warming ocean increases surface pCO2 through changes in marine ecosystem composition. Glob. Biogeochem. Cycle 2013, 27, 1214–1225. [Google Scholar] [CrossRef]
- Armstrong McKay, D.I.; Cornell, S.E.; Richardson, K.; Rockström, J. Resolving ecological feedbacks on the ocean carbon sink in Earth system models. Earth Syst. Dynam. 2021, 12, 797–818. [Google Scholar] [CrossRef]
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef]
- Pomeroy, L.R.; Wiebe, W.J. Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria. Aquat. Microb. Ecol. 2001, 23, 187–204. [Google Scholar] [CrossRef]
- Brewer, P.G.; Peltzer, E.T. Depth perception: The need to report ocean biogeochemical rates as functions of temperature, not depth. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2017, 375, 20160319. [Google Scholar] [CrossRef] [PubMed]
- Gabric, A.J.; Gregg, W.; Najarr, R.; Erickson, D.; Matrai, P. Modelling the biogeochemical cycle of dimethylsulphide in the upper ocean. Chemosphere Glob. Chang. Sci. 2001, 3, 377–392. [Google Scholar] [CrossRef]
- Moloney, C.L.; Field, J.G. Size-based dynamics of plankton food webs. I. A simulation model of carbon and nitrogen flows. J. Plankton Res. 1991, 13, 1003–1038. [Google Scholar] [CrossRef]
- Gabric, A.J.; Matrai, P.; Vernet, M. Modelling the production of dimethylsulphide during the vernal bloom in the Barents Sea. Tellus B 1999, 51, 919–938. [Google Scholar] [CrossRef]
- Qu, B.; Gabric, A.J. Using genetic algorithms to calibrate a dimethylsulfide production model in the Arctic Ocean. Chin. J. Oceanol. Limnol. 2010, 28, 573–582. [Google Scholar] [CrossRef]
- Gabric, A.J.; Whetton, P.; Cropp, R. Dimethylsulphide production in the subantarctic Southern Ocean under enhanced greenhouse conditions. Tellus B 2001, 53, 273–287. [Google Scholar] [CrossRef]
- Watts, M.; Bigg, G. Modelling the nitrogen cycle and DMS production in Lagrangian experiments in the North Atlantic. Deep.-Sea Res. II 2001, 48, 1019–1042. [Google Scholar] [CrossRef]
- Qu, B.; Zhao, L.; Gabric, A.J. Simulating the sea-to-air flux of dimethylsulfide sea in the eastern China marginal seas. J. Mar. Syst. 2020, 212, 103450. [Google Scholar] [CrossRef]
- Qu, B.; Gabric, A.J.; Zeng, M.; Lu, Z. Dimethylsulfide model calibration in the Barents Sea using a genetic algorithm and neural network. Environ. Chem. 2015, 13, 413–424. [Google Scholar] [CrossRef]
- Halloran, P.R.; Bell, T.G.; Totterdell, I.J. Can we trust empirical marine DMS parameterisations within projections of future climate? Biogeosciences 2010, 7, 1645–1656. [Google Scholar] [CrossRef]
- Hind, A.J.; Rauschenberg, C.D.; Johnson, J.E.; Yang, M.; Matrai, P.A. The use of algorithms to predict surface seawater dimethyl sulphide concentrations in the SE Pacific, a region of steep gradients in primary productivity, biomass and mixed layer depth. Biogeosciences 2011, 8, 1–16. [Google Scholar] [CrossRef]
- Anderson, T.R.; Spall, S.A.; Yool, A.; Cipollini, P.; Challenor, P.G.; Fasham, M.J.R. Global fields of sea surface dimethylsulfide predicted from chlorophyll, nutrients and light. J. Mar. Syst. 2001, 30, 1–20. [Google Scholar] [CrossRef]
- Galí, M.; Levasseur, M.; Devred, E.; Simó, R.; Babin, M. Sea-surface dimethylsulfide (DMS) concentration from satellite data at global and regional scales. Biogeosciences 2018, 15, 3497–3519. [Google Scholar] [CrossRef]
- Herr, A.E.; Kiene, R.P.; Dacey, J.W.H.; Tortell, P.D. Patterns and drivers of dimethylsulfide concentration in the northeast subarctic Pacific across multiple spatial and temporal scales. Biogeosciences 2019, 16, 1729–1754. [Google Scholar] [CrossRef]
- McNabb, B.J.; Tortell, P.D. Improved prediction of dimethyl sulfide (DMS) distributions in the northeast subarctic Pacific using machine-learning algorithms. Biogeosciences 2022, 19, 1705–1721. [Google Scholar] [CrossRef]
- Tesdal, J.-E.; Christian, J.R.; Monahan, A.H.; von Salzen, K. Evaluation of diverse approaches for estimating sea-surface DMS concentration and air–sea exchange at global scale. Environ. Chem. 2016, 13, 390–412. [Google Scholar] [CrossRef]
- Gabric, A.J.; Cropp, R.A.; Hirst, A.C.; Marchant, H.J. The response of dimethylsulphide production to simulated warming in the eastern Antarctic Southern Ocean. Tellus B 2003, 55, 966–981. [Google Scholar] [CrossRef]
- Gabric, A.J.; Qu, B.; Matrai, P.; Hirst, A.C. The simulated response of dimethylsulfide production in the Arctic Ocean to global warming. Tellus B 2005, 57, 391–403. [Google Scholar] [CrossRef]
- Bopp, L.; Aumont, O.; Belviso, S.; Monfray, P. Potential impact of climate change on marine dimethyl sulfide emissions. Tellus Ser. B-Chem. Phys. Meteorol. 2003, 55, 11–22. [Google Scholar] [CrossRef]
- Wang, S.; Maltrud, M.; Elliott, S.; Cameron-Smith, P.; Jonko, A. Influence of dimethyl sulfide on the carbon cycle and biological production. Biogeochemistry 2018, 138, 49–68. [Google Scholar] [CrossRef]
- Hopkins, F.E.; Suntharalingam, P.; Gehlen, M.; Andrews, O.; Archer, S.D.; Bopp, L.; Buitenhuis, E.; Dadou, I.; Duce, R.; Goris, N.; et al. The impacts of ocean acidification on marine trace gases and the implications for atmospheric chemistry and climate. Proc. R. Soc. A Math. Phys. Eng. Sci. 2020, 476, 20190769. [Google Scholar] [CrossRef]
- Taucher, J.; Oschlies, A. Can we predict the direction of marine primary production change under global warming? Geophys. Res. Lett. 2011, 38. [Google Scholar] [CrossRef]
- Laufkötter, C.; Vogt, M.; Gruber, N.; Aita-Noguchi, M.; Aumont, O.; Bopp, L.; Buitenhuis, E.; Doney, S.C.; Dunne, J.; Hashioka, T.; et al. Drivers and uncertainties of future global marine primary production in marine ecosystem models. Biogeosciences 2015, 12, 6955–6984. [Google Scholar] [CrossRef]
- Friedlingstein, P.; Jones, M.W.; O’Sullivan, M.; Andrew, R.M.; Bakker, D.C.E.; Hauck, J.; Le Quéré, C.; Peters, G.P.; Peters, W.; Pongratz, J.; et al. Global Carbon Budget 2021. Earth Syst. Sci. Data 2022, 14, 1917–2005. [Google Scholar] [CrossRef]
- Avgoustidi, V.; Nightingale, P.D.; Joint, I.; Steinke, M.; Turner, S.M.; Hopkins, F.E.; Liss, P.S. Decreased marine dimethyl sulfide production under elevated CO2 levels in mesocosm and in vitro studies. Environ. Chem. 2012, 9, 399–404. [Google Scholar] [CrossRef]
- Spielmeyer, A.; Pohnert, G. Influence of temperature and elevated carbon dioxide on the production of dimethylsulfoniopropionate and glycine betaine by marine phytoplankton. Mar. Environ. Res. 2012, 73, 62–69. [Google Scholar] [CrossRef]
- Arnold, H.E.; Kerrison, P.; Steinke, M. Interacting effects of ocean acidification and warming on growth and DMS-production in the haptophyte coccolithophore Emiliania huxleyi. Glob. Chang. Biol. 2013, 19, 1007–1016. [Google Scholar] [CrossRef]
- Park, K.-T.; Lee, K.; Shin, K.; Yang, E.J.; Hyun, B.; Kim, J.-M.; Noh, J.H.; Kim, M.; Kong, B.; Choi, D.H.; et al. Direct Linkage between Dimethyl Sulfide Production and Microzooplankton Grazing, Resulting from Prey Composition Change under High Partial Pressure of Carbon Dioxide Conditions. Environ. Sci. Technol. 2014, 48, 4750–4756. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, F.E.; Archer, S.D. Consistent increase in dimethyl sulfide (DMS) in response to high CO2 in five shipboard bioassays from contrasting NW European waters. Biogeosciences 2014, 11, 4925–4940. [Google Scholar] [CrossRef]
- Li, P.-F.; Yang, G.-P.; Liu, C.-Y. Combined effects of elevated temperature and pCO2 on the production of DMSP and DMS in the culture of Amphidinium carterae. J. Appl. Phycol. 2020, 32, 1063–1074. [Google Scholar] [CrossRef]
- Saint-Macary, A.D.; Barr, N.; Armstrong, E.; Safi, K.; Marriner, A.; Gall, M.; McComb, K.; Dillingham, P.W.; Law, C.S. The Influence of Ocean Acidification and Warming on DMSP & DMS in New Zealand Coastal Water. Atmosphere 2021, 12, 181. [Google Scholar]
- Bénard, R.; Levasseur, M.; Scarratt, M.; Michaud, S.; Starr, M.; Mucci, A.; Ferreyra, G.; Gosselin, M.; Tremblay, J.É.; Lizotte, M.; et al. Contrasting effects of acidification and warming on dimethylsulfide concentrations during a temperate estuarine fall bloom mesocosm experiment. Biogeosciences 2019, 16, 1167–1185. [Google Scholar] [CrossRef]
- Benard, R.; Lizotte, M.; Levasseur, M.; Scarratt, M.; Michaud, S.; Starr, M.; Tremblay, J.E.; Kiene, R.P.; Kameyama, S. Impact of anthropogenic pH perturbation on dimethyl sulfide cycling: A peek into the microbial black box. Elem.-Sci. Anthr. 2021, 9, 00043. [Google Scholar] [CrossRef]
- Hopkins, F.E.; Turner, S.M.; Nightingale, P.D.; Steinke, M.; Bakker, D.; Liss, P.S. Ocean acidification and marine trace gas emissions. Proc. Natl. Acad. Sci. USA 2010, 107, 760–765. [Google Scholar] [CrossRef]
- Dix, M.; Bi, D.; Dobrohotoff, P.; Fiedler, R.; Harman, I.; Law, R.; Mackallah, C.; Marsland, S.; O’Farrell, S.; Rashid, H.; et al. CSIRO-ARCCSS ACCESS-CM2 Model Output Prepared for CMIP6. Earth System Grid Federation. 2019. Available online: https://www.wdc-climate.de/ui/cmip6?input=CMIP6.ScenarioMIP.CSIRO-ARCCSS.ACCESS-CM2.ssp245 (accessed on 26 July 2022). [CrossRef]
- Krasting, J.P.; John, J.G.; Blanton, C.; McHugh, C.; Nikonov, S.; Radhakrishnan, A.; Rand, K.; Zadeh, N.T.; Balaji, V.; Durachta, J.; et al. NOAA-GFDL GFDL-ESM4 Model Output Prepared for CMIP6. Earth System Grid Federation. 2018. Available online: https://www.wdc-climate.de/ui/cmip6?input=CMIP6.CMIP.NOAA-GFDL.GFDL-ESM4 (accessed on 26 July 2022). [CrossRef]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef]
- Fricko, O.; Havlik, P.; Rogelj, J.; Klimont, Z.; Gusti, M.; Johnson, N.; Kolp, P.; Strubegger, M.; Valin, H.; Amann, M. The marker quantification of the Shared Socioeconomic Pathway 2: A middle-of-the-road scenario for the 21st century. Glob. Environ. Chang. 2017, 42, 251–267. [Google Scholar] [CrossRef]
- Kriegler, E.; Bauer, N.; Popp, A.; Humpenöder, F.; Leimbach, M.; Strefler, J.; Baumstark, L.; Bodirsky, B.L.; Hilaire, J.; Klein, D. Fossil-fueled development (SSP5): An energy and resource intensive scenario for the 21st century. Glob. Environ. Chang. 2017, 42, 297–315. [Google Scholar] [CrossRef]
- IPPC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; Core Writing Team; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Le Quéré, C.; Andrew, R.M.; Friedlingstein, P.; Sitch, S.; Hauck, J.; Pongratz, J.; Pickers, P.A.; Korsbakken, J.I.; Peters, G.P.; Canadell, J.G.; et al. Global Carbon Budget. Earth Syst. Sci. Data 2018, 10, 2141–2194. [Google Scholar] [CrossRef]
- Gabric, A.; Matrai, P.; Jones, G.; Middleton, J. The nexus between sea ice and polar emissions of marine biogenic aerosols. Bull. Am. Meteorol. Soc. 2018, 99, 61–82. [Google Scholar] [CrossRef]
- Asher, E.C.; Dacey, J.W.H.; Stukel, M.; Long, M.C.; Tortell, P.D. Processes driving seasonal variability in DMS, DMSP, and DMSO concentrations and turnover in coastal Antarctic waters. Limnol. Oceanogr. 2017, 62, 104–124. [Google Scholar] [CrossRef]
- Doney, S.C.; Ruckelshaus, M.; Emmett Duffy, J.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate Change Impacts on Marine Ecosystems. Annu. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef]
- Kirchman, D.L.; Morán, X.A.G.; Ducklow, H. Microbial growth in the polar oceans—Role of temperature and potential impact of climate change. Nat. Rev. Microbiol. 2009, 7, 451–459. [Google Scholar] [CrossRef]
- Kim, H.; Ducklow, H.W. A Decadal (2002–2014) Analysis for Dynamics of Heterotrophic Bacteria in an Antarctic Coastal Ecosystem: Variability and Physical and Biogeochemical Forcings. Front. Mar. Sci. 2016, 3, 214. [Google Scholar] [CrossRef]
- Orr, J.C.; Fabry, V.J.; Aumont, O.; Bopp, L.; Doney, S.C.; Feely, R.A.; Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681–686. [Google Scholar] [CrossRef]
- Shadwick, E.H.; Trull, T.W.; Thomas, H.; Gibson, J.A.E. Vulnerability of Polar Oceans to Anthropogenic Acidification: Comparison of Arctic and Antarctic Seasonal Cycles. Sci. Rep. 2013, 3, 2339. [Google Scholar] [CrossRef]
- Tynan, E.; Clarke, J.S.; Humphreys, M.P.; Ribas-Ribas, M.; Esposito, M.; Rérolle, V.M.C.; Schlosser, C.; Thorpe, S.E.; Tyrrell, T.; Achterberg, E.P. Physical and biogeochemical controls on the variability in surface pH and calcium carbonate saturation states in the Atlantic sectors of the Arctic and Southern Oceans. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2016, 127, 7–27. [Google Scholar] [CrossRef]
- Hussherr, R.; Levasseur, M.; Lizotte, M.; Tremblay, J.É.; Mol, J.; Thomas, H.; Gosselin, M.; Starr, M.; Miller, L.A.; Jarniková, T.; et al. Impact of ocean acidification on Arctic phytoplankton blooms and dimethyl sulfide concentration under simulated ice-free and under-ice conditions. Biogeosciences 2017, 14, 2407–2427. [Google Scholar] [CrossRef]
- Archer, S.D.; Kimmance, S.A.; Stephens, J.A.; Hopkins, F.E.; Bellerby, R.G.J.; Schulz, K.G.; Piontek, J.; Engel, A. Contrasting responses of DMS and DMSP to ocean acidification in Arctic waters. Biogeosciences 2013, 10, 1893–1908. [Google Scholar] [CrossRef]
- Hopkins, F.E.; Nightingale, P.D.; Stephens, J.A.; Moore, C.M.; Richier, S.; Cripps, G.L.; Archer, S.D. Dimethylsulfide (DMS) production in polar oceans may be resilient to ocean acidification. Biogeosci. Discuss 2018, 10, 1–42. [Google Scholar]
- Hopkins, F.E.; Nightingale, P.D.; Stephens, J.A.; Moore, C.M.; Richier, S.; Cripps, G.L.; Archer, S.D. A meta-analysis of microcosm experiments shows that dimethyl sulfide (DMS) production in polar waters is insensitive to ocean acidification. Biogeosciences 2020, 17, 163–186. [Google Scholar] [CrossRef]
- Baker, A.C.; Glynn, P.W.; Riegl, B. Climate change and coral reef bleaching: An ecological assessment of long-term impacts, recovery trends and future outlook. Estuar. Coast. Shelf Sci. 2008, 80, 435–471. [Google Scholar] [CrossRef]
- Heron, S.F.; Maynard, J.A.; Van Hooidonk, R.; Eakin, C.M. Warming trends and bleaching stress of the world’s coral reefs 1985–2012. Sci. Rep. 2016, 6, 38402. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Chase, T.J.; Dietzel, A.; Hill, T.; Hoey, A.S.; Hoogenboom, M.O.; Jacobson, M. Global warming impairs stock–recruitment dynamics of corals. Nature 2019, 568, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.F.; De’Ath, G.; Fabricius, K.E.; Lough, J.M. Declining coral calcification in massive Porites in two nearshore regions of the northern Great Barrier Reef. Glob. Chang. Biol. 2008, 14, 529–538. [Google Scholar] [CrossRef]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. Insights into the coral microbiome: Underpinning the health and resilience of reef ecosystems. Annu. Rev. Microbiol. 2016, 70, 317–340. [Google Scholar] [CrossRef]
- Lesser, M.P. Coral bleaching: Causes and mechanisms. In Coral Reefs: An Ecosystem in Transition; Springer: Berlin/Heidelberg, Germany, 2011; pp. 405–419. [Google Scholar]
- Berkelmans, R. Bleaching and mortality thresholds: How much is too much? In Coral Bleaching. Ecological Studies; van Oppen, M.J.H., Lough, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar] [CrossRef]
- Burke, L.; Reytar, K.; Spalding, M.; Perry, A. Reefs at Risk Revisited; World Resources Institute: Washington, DC, USA, 2011. [Google Scholar]
- Ainsworth, T.D.; Heron, S.F.; Ortiz, J.C.; Mumby, P.J.; Grech, A.; Ogawa, D.; Eakin, C.M.; Leggat, W. Climate change disables coral bleaching protection on the Great Barrier Reef. Science 2016, 352, 338–342. [Google Scholar] [CrossRef]
- Dixon, A.M.; Forster, P.M.; Heron, S.F.; Stoner, A.M.K.; Beger, M. Future loss of local-scale thermal refugia in coral reef ecosystems. PLoS Clim. 2022, 1, e0000004. [Google Scholar] [CrossRef]
- Jackson, R.L.; Gabric, A.J.; Matrai, P.A.; Woodhouse, M.T.; Cropp, R.; Jones, G.B.; Deschaseaux, E.S.M.; Omori, Y.; McParland, E.L.; Swan, H.B. Parameterizing the impact of seawater temperature and irradiance on dimethylsulfide (DMS) in the Great Barrier Reef and the contribution of coral reefs to the global sulfur cycle. J. Geophys. Res. Ocean. 2021, 126, e012020JC016783. [Google Scholar] [CrossRef]
- Jackson, R.L.; Gabric, A.J.; Woodhouse, M.T.; Swan, H.B.; Jones, G.B.; Cropp, R.; Deschaseaux, E.S.M. Coral reef emissions of atmospheric dimethylsulfide and the influence on marine aerosols in the southern Great Barrier Reef, Australia. J. Geophys. Res. Atmos. 2020, 125, e032019JD031837. [Google Scholar] [CrossRef]
- Swan, H.B.; Jones, G.B.; Deschaseaux, E.S.M.; Eyre, B.D. Coral reef origins of atmospheric dimethylsulfide at Heron Island, southern Great Barrier Reef, Australia. Biogeosciences 2017, 14, 229–239. [Google Scholar] [CrossRef]
- Jones, G.B.; Curran, M.; Broadbent, A.; King, S.; Fischer, E.; Jones, R. Factors affecting the cycling of dimethylsulfide and dimethylsulfoniopropionate in coral reef waters of the Great Barrier Reef. Environ. Chem. 2007, 4, 310–322. [Google Scholar] [CrossRef]
- Jones, G.B.; Fischer, E.; Deschaseaux, E.S.M.; Harrison, P.L. The effect of coral bleaching on the cellular concentration of dimethylsulphoniopropionate in reef corals. J. Exp. Mar. Biol. Ecol. 2014, 460, 19–31. [Google Scholar] [CrossRef]
- Raina, J.-B.; Tapiolas, D.; Willis, B.L.; Bourne, D.G. Coral-associated bacteria and their role in the biogeochemical cycling of sulfur. Appl. Environ. Microbiol. 2009, 75, 3492–3501. [Google Scholar] [CrossRef]
- Deschaseaux, E.S.M.; Jones, G.B.; Deseo, M.A.; Shepherd, K.M.; Kiene, R.P.; Swan, H.B.; Harrison, P.L.; Eyre, B.D. Effects of environmental factors on dimethylated sulfur compounds and their potential role in the antioxidant system of the coral holobiont. Limnol. Oceanogr. 2014, 59, 758–768. [Google Scholar] [CrossRef]
- Gardner, S.G.; Nielsen, D.A.; Laczka, O.; Shimmon, R.; Beltran, V.H.; Ralph, P.J.; Petrou, K. Dimethylsulfoniopropionate, superoxide dismutase and glutathione as stress response indicators in three corals under short-term hyposalinity stress. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152418. [Google Scholar] [CrossRef]
- Jones, R.J.; Hoegh-Guldberg, O.; Larkum, A.W.D.; Schreiber, U. Temperature-induced bleaching of corals begins with impairment of the CO2 fixation mechanism in zooxanthellae. Plant Cell Environ. 2002, 21, 1219–1230. [Google Scholar] [CrossRef]
- Lesser, M.P.; Stochaj, W.R.; Tapley, D.W.; Shick, J.M. Bleaching in coral reef anthozoans: Effects of irradiance, ultraviolet radiation, and temperature on the activities of protective enzymes against active oxygen. Coral Reefs 1990, 8, 225–232. [Google Scholar] [CrossRef]
- Bay, L.K.; Doyle, J.; Logan, M.; Berkelmans, R. Recovery from bleaching is mediated by threshold densities of background thermo-tolerant symbiont types in a reef-building coral. R. Soc. Open Sci. 2016, 3, 160322. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, D.A.; Petrou, K.; Gates, R.D. Coral bleaching from a single cell perspective. ISME J. 2018, 12, 1558–1567. [Google Scholar] [CrossRef] [PubMed]
- Berkelmans, R.; Van Oppen, M.J.H. The role of zooxanthellae in the thermal tolerance of corals: A ‘nugget of hope’for coral reefs in an era of climate change. Proc. R. Soc. B Biol. Sci. 2006, 273, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.B.; King, S. Dimethylsulphoniopropionate (DMSP) as an indicator of bleaching tolerance in scleractinian corals. J. Mar. Sci. Eng. 2015, 3, 444–465. [Google Scholar] [CrossRef]
- Deschaseaux, E.S.M.; Beltran, V.H.; Jones, G.B.; Deseo, M.A.; Swan, H.B.; Harrison, P.L.; Eyre, B.D. Comparative response of DMS and DMSP concentrations in Symbiodinium clades C1 and D1 under thermal stress. J. Exp. Mar. Biol. Ecol. 2014, 459, 181–189. [Google Scholar] [CrossRef]
- Klueter, A.; Trapani, J.; Archer, F.I.; McIlroy, S.E.; Coffroth, M.A. Comparative growth rates of cultured marine dinoflagellates in the genus Symbiodinium and the effects of temperature and light. PLoS ONE 2017, 12, e0187707. [Google Scholar] [CrossRef]
- Jackson, R.; Gabric, A.; Cropp, R. Effects of ocean warming and coral bleaching on aerosol emissions in the Great Barrier Reef, Australia. Sci. Rep. 2018, 8, 14048. [Google Scholar] [CrossRef]
- Fischer, E.; Jones, G. Atmospheric dimethysulphide production from corals in the Great Barrier Reef and links to solar radiation, climate and coral bleaching. Biogeochemistry 2012, 110, 31–46. [Google Scholar] [CrossRef]
- Orr, J.C. Recent and future changes in ocean carbonate chemistry. Ocean. Acidif. 2011, 1, 41–66. [Google Scholar]
- Brodie, J.E.; Devlin, M.; Haynes, D.; Waterhouse, J. Assessment of the eutrophication status of the Great Barrier Reef lagoon (Australia). Biogeochemistry 2011, 106, 281–302. [Google Scholar] [CrossRef]
- Howarth, R.; Chan, F.; Conley, D.J.; Garnier, J.; Doney, S.C.; Marino, R.; Billen, G. Coupled biogeochemical cycles: Eutrophication and hypoxia in temperate estuaries and coastal marine ecosystems. Front. Ecol. Environ. 2011, 9, 18–26. [Google Scholar] [CrossRef]
- McKergow, L.A.; Prosser, I.P.; Hughes, A.O.; Brodie, J. Sources of sediment to the Great Barrier Reef world heritage area. Mar. Pollut. Bull. 2005, 51, 200–211. [Google Scholar] [CrossRef]
- Gabric, A.J.; Bell, P.R. Review of the effects of non-point nutrient loading on coastal ecosystems. Mar. Freshw. Res. 1993, 44, 261–283. [Google Scholar] [CrossRef]
- McCook, L.J.; Diaz-Pulido, G. The fate of bleached corals: Patterns and dynamics of algal recruitment. Mar. Ecol. Prog. Ser. 2002, 232, 115–128. [Google Scholar] [CrossRef]
- De’ath, G.; Fabricius, K. Water quality as a regional driver of coral biodiversity and macroalgae on the Great Barrier Reef. Ecol. Appl. 2010, 20, 840–850. [Google Scholar] [CrossRef]
Figure 1.
Simplified schematic representation of the dimethylsufide (DMS) and dimethylsulfoniopropionate (DMSP) cycles. Red-colored processes indicate those that are mediated by heterotrophic bacteria. DMSO: dimethylsulfoxide.
Figure 1.
Simplified schematic representation of the dimethylsufide (DMS) and dimethylsulfoniopropionate (DMSP) cycles. Red-colored processes indicate those that are mediated by heterotrophic bacteria. DMSO: dimethylsulfoxide.
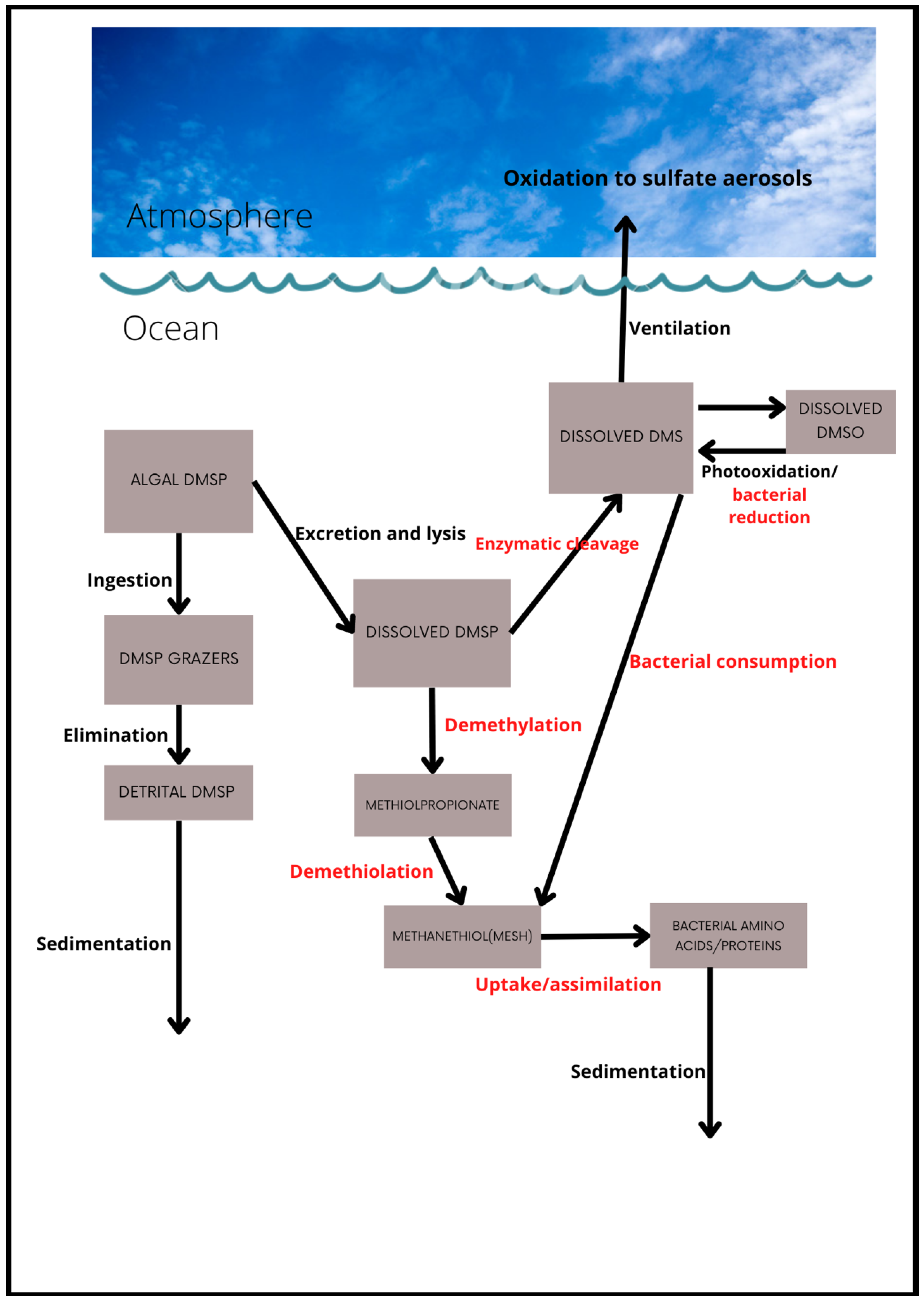
Figure 2.
(a) Present-day annual mean sea surface temperature (SST) and the change in SST by the end of this century, as simulated by CMIP6 climate models for climate under two emission scenarios: (b) SSP2-4.5 and (c) SSP5-8.5.
Figure 2.
(a) Present-day annual mean sea surface temperature (SST) and the change in SST by the end of this century, as simulated by CMIP6 climate models for climate under two emission scenarios: (b) SSP2-4.5 and (c) SSP5-8.5.

Figure 3.
(a) Present-day annual mean mixed layer depth (MLD) and the change in MLD by the end of this century, as simulated for climate under two emission scenarios: (b) SSP2-4.5 and (c) SSP5-8.5.
Figure 3.
(a) Present-day annual mean mixed layer depth (MLD) and the change in MLD by the end of this century, as simulated for climate under two emission scenarios: (b) SSP2-4.5 and (c) SSP5-8.5.
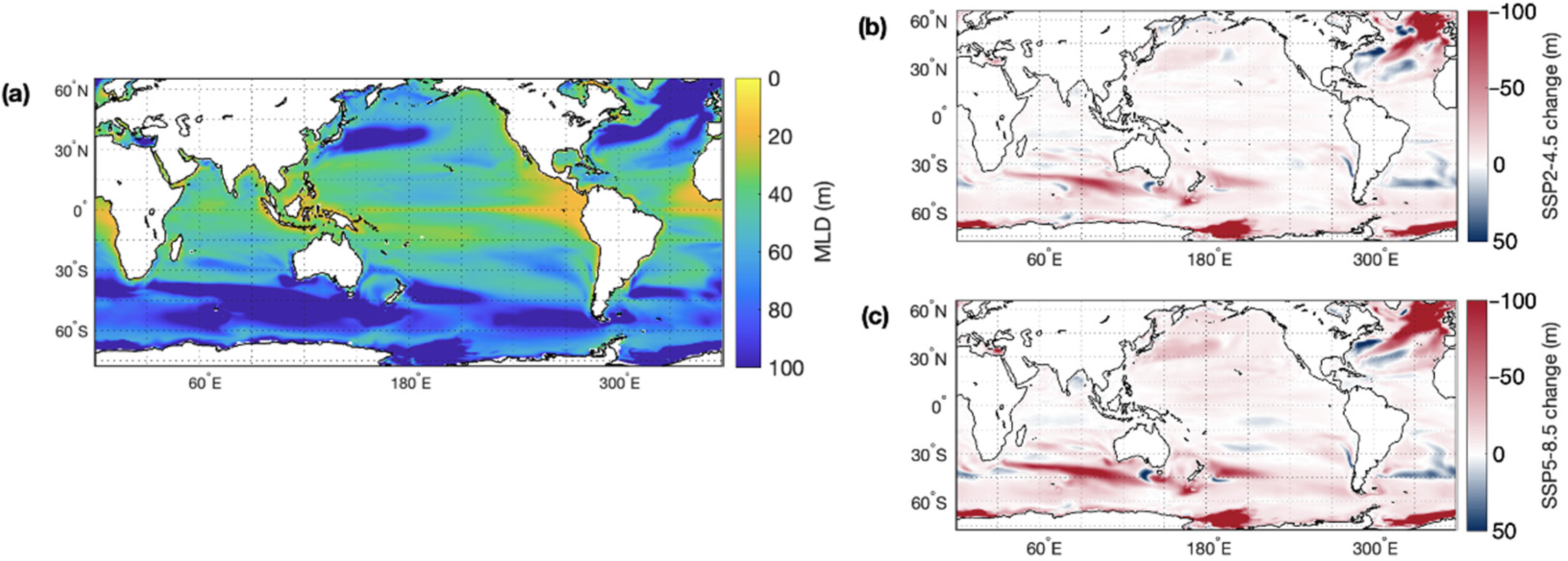
Figure 4.
(a) Annual mean pH and the change in pH by the end of this century, as simulated for climate under two emission scenarios: (b) SSP2-4.5 and (c) SSP5-8.5.
Figure 4.
(a) Annual mean pH and the change in pH by the end of this century, as simulated for climate under two emission scenarios: (b) SSP2-4.5 and (c) SSP5-8.5.
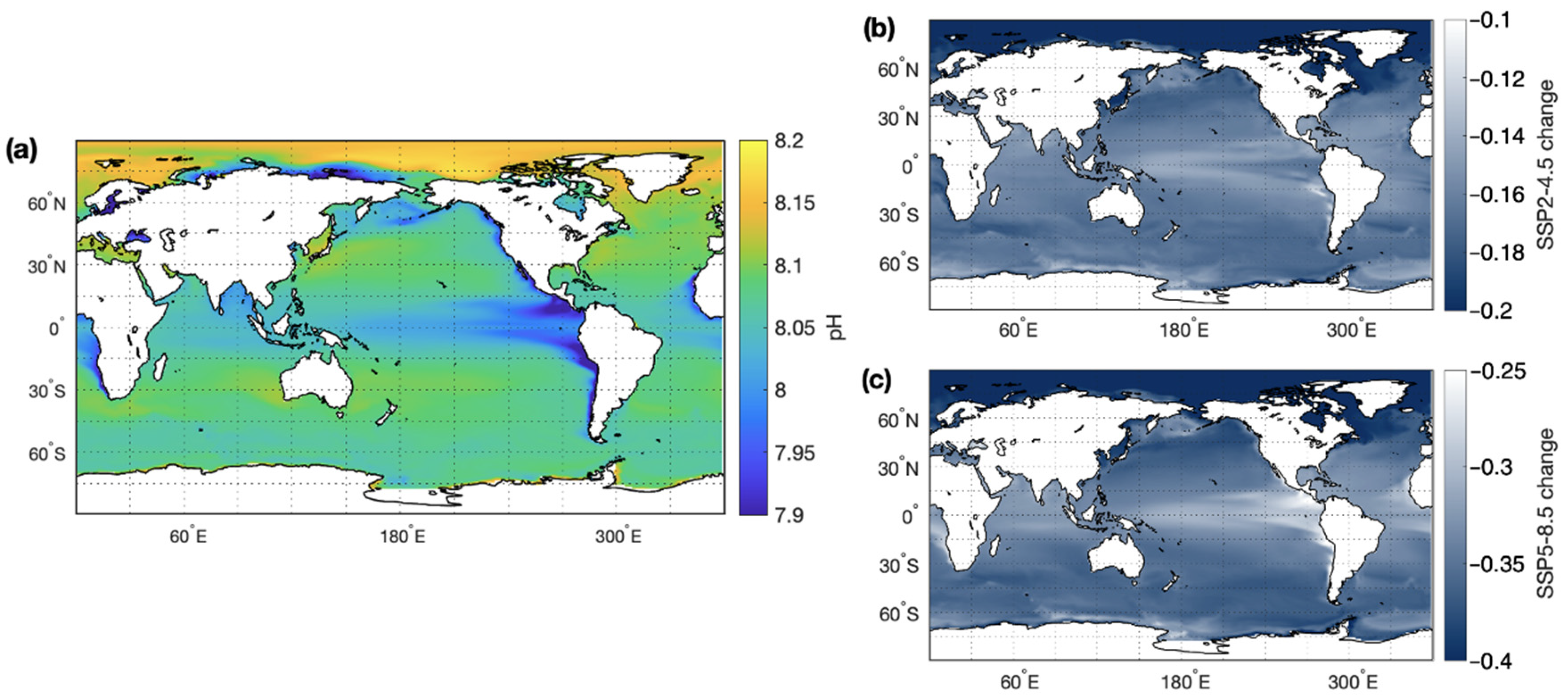
Figure 5.
Climatology (±2 SE) of (a) SST and (b) MLD for a contemporary (black) and end-of-century climate modeled under two emission scenarios: SSP2-4.5 (blue) and SSP5-8.5 (red) scenarios for the tropical ocean (20° N–20° S).
Figure 5.
Climatology (±2 SE) of (a) SST and (b) MLD for a contemporary (black) and end-of-century climate modeled under two emission scenarios: SSP2-4.5 (blue) and SSP5-8.5 (red) scenarios for the tropical ocean (20° N–20° S).
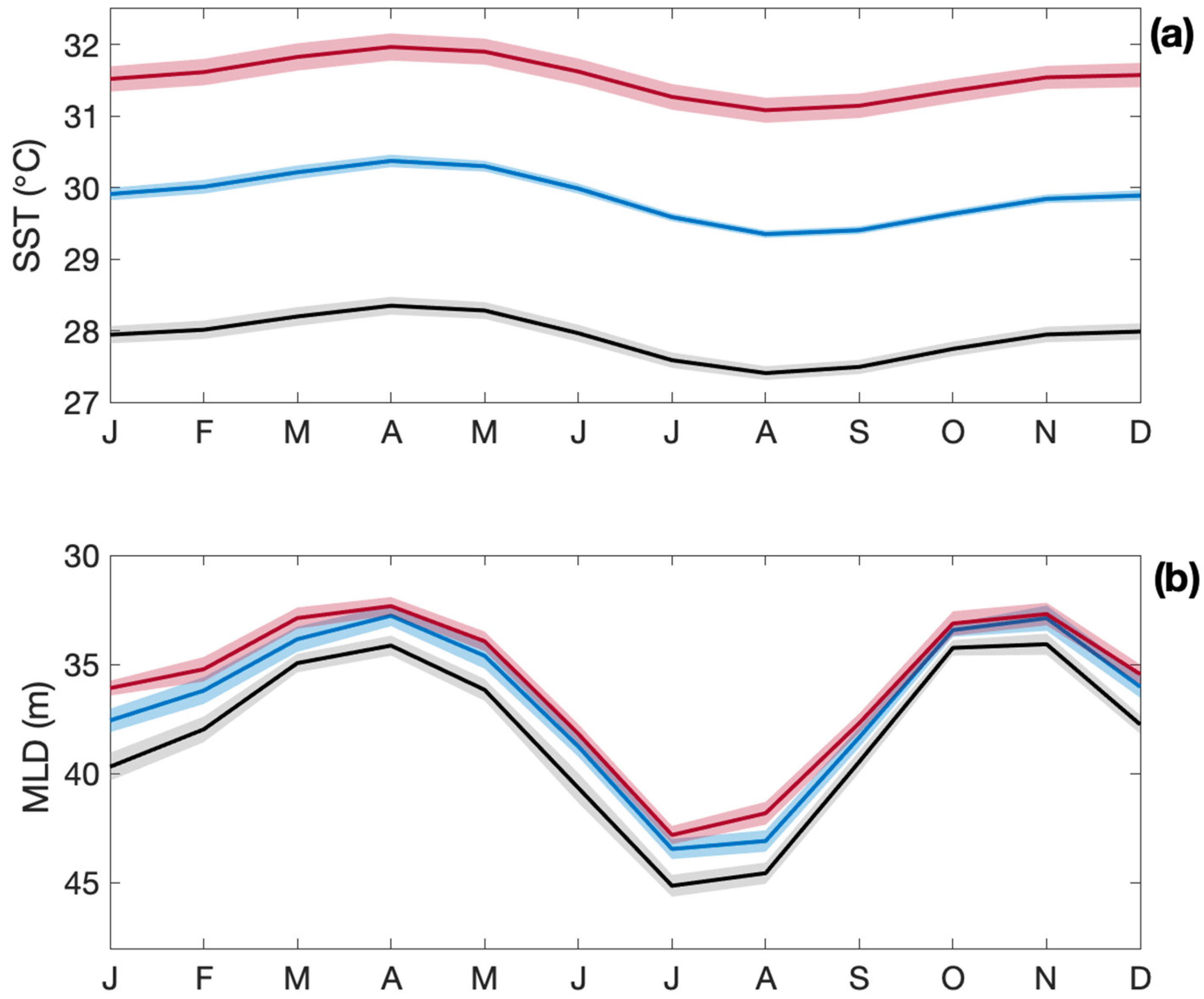
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jackson, R.; Gabric, A. Climate Change Impacts on the Marine Cycling of Biogenic Sulfur: A Review. Microorganisms 2022, 10, 1581. https://doi.org/10.3390/microorganisms10081581
AMA Style
Jackson R, Gabric A. Climate Change Impacts on the Marine Cycling of Biogenic Sulfur: A Review. Microorganisms. 2022; 10(8):1581. https://doi.org/10.3390/microorganisms10081581
Chicago/Turabian StyleJackson, Rebecca, and Albert Gabric. 2022. "Climate Change Impacts on the Marine Cycling of Biogenic Sulfur: A Review" Microorganisms 10, no. 8: 1581. https://doi.org/10.3390/microorganisms10081581
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.